Home — Essay Samples — Science — Experiment — Introduction to Mendelian Genetics

Introduction to Mendelian Genetics
- Categories: Experiment
About this sample

Words: 887 |
Published: Apr 11, 2019
Words: 887 | Pages: 2 | 5 min read
Materials and Methods:
- Mung Bean Plants
Discussion:

Cite this Essay
Let us write you an essay from scratch
- 450+ experts on 30 subjects ready to help
- Custom essay delivered in as few as 3 hours
Get high-quality help

Dr. Karlyna PhD
Verified writer
- Expert in: Science

+ 120 experts online
By clicking “Check Writers’ Offers”, you agree to our terms of service and privacy policy . We’ll occasionally send you promo and account related email
No need to pay just yet!
Related Essays
1 pages / 678 words
1 pages / 441 words
2 pages / 790 words
1 pages / 411 words
Remember! This is just a sample.
You can get your custom paper by one of our expert writers.
121 writers online
Still can’t find what you need?
Browse our vast selection of original essay samples, each expertly formatted and styled
Related Essays on Experiment
The Gummy Bear Lab is a popular experiment in chemistry classrooms, designed to illustrate principles such as osmosis, diffusion, and chemical reactions. This lab typically involves immersing gummy bears in various solutions [...]
Since childhood, I have always been fascinated by the human body and the intricate mechanisms that govern its functioning. This fascination has been the driving force behind my ambition to pursue a career as a doctor. In this [...]
The Tuskegee Experiment, also known as the Tuskegee Study of Untreated Syphilis in the Negro Male, is considered one of the most notorious instances of unethical conduct in the history of medical research. The study, which began [...]
Addiction remains one of the most complex and pervasive issues facing modern society. The study of addiction, particularly through controlled experiments, offers valuable insights into the mechanisms behind addictive behaviors [...]
On April 4, 1968 Dr. Martin Luther King, Jr was assassinated and many people were shocked because of this. “A Class Divided” which is a video tells as what happened on the day after this sad day. Because of this, the third [...]
To observe the constituents and structure of metals and their alloys by means of an optical microscope. This is about to examine the microstructure of steel and observe the constituents and structure of it. Microstructure [...]
Related Topics
By clicking “Send”, you agree to our Terms of service and Privacy statement . We will occasionally send you account related emails.
Where do you want us to send this sample?
By clicking “Continue”, you agree to our terms of service and privacy policy.
Be careful. This essay is not unique
This essay was donated by a student and is likely to have been used and submitted before
Download this Sample
Free samples may contain mistakes and not unique parts
Sorry, we could not paraphrase this essay. Our professional writers can rewrite it and get you a unique paper.
Please check your inbox.
We can write you a custom essay that will follow your exact instructions and meet the deadlines. Let's fix your grades together!
Get Your Personalized Essay in 3 Hours or Less!
We use cookies to personalyze your web-site experience. By continuing we’ll assume you board with our cookie policy .
- Instructions Followed To The Letter
- Deadlines Met At Every Stage
- Unique And Plagiarism Free
This page has been archived and is no longer updated
Gregor Mendel and the Principles of Inheritance
Traits are passed down in families in different patterns. Pedigrees can illustrate these patterns by following the history of specific characteristics, or phenotypes, as they appear in a family. For example, the pedigree in Figure 1 shows a family in which a grandmother (generation I) has passed down a characteristic (shown in solid red) through the family tree. The inheritance pattern of this characteristic is considered dominant , because it is observable in every generation. Thus, every individual who carries the genetic code for this characteristic will show evidence of the characteristic. In contrast, Figure 2 shows a different pattern of inheritance, in which a characteristic disappears in one generation, only to reappear in a subsequent one. This pattern of inheritance, in which the parents do not show the phenotype but some of the children do, is considered recessive . But where did our knowledge of dominance and recessivity first come from?
Gregor Mendel’s Courage and Persistence
Mendel was curious about how traits were transferred from one generation to the next, so he set out to understand the principles of heredity in the mid-1860s. Peas were a good model system, because he could easily control their fertilization by transferring pollen with a small paintbrush. This pollen could come from the same flower (self-fertilization), or it could come from another plant's flowers (cross-fertilization). First, Mendel observed plant forms and their offspring for two years as they self-fertilized, or "selfed," and ensured that their outward, measurable characteristics remained constant in each generation. During this time, Mendel observed seven different characteristics in the pea plants, and each of these characteristics had two forms (Figure 3). The characteristics included height (tall or short), pod shape (inflated or constricted), seed shape (smooth or winkled), pea color (green or yellow), and so on. In the years Mendel spent letting the plants self, he verified the purity of his plants by confirming, for example, that tall plants had only tall children and grandchildren and so forth. Because the seven pea plant characteristics tracked by Mendel were consistent in generation after generation of self-fertilization, these parental lines of peas could be considered pure-breeders (or, in modern terminology, homozygous for the traits of interest). Mendel and his assistants eventually developed 22 varieties of pea plants with combinations of these consistent characteristics.
Mendel not only crossed pure-breeding parents, but he also crossed hybrid generations and crossed the hybrid progeny back to both parental lines. These crosses (which, in modern terminology, are referred to as F 1 , F 1 reciprocal, F 2 , B 1 , and B 2 ) are the classic crosses to generate genetically hybrid generations.
Understanding Dominant Traits
Understanding recessive traits.
When conducting his experiments, Mendel designated the two pure-breeding parental generations involved in a particular cross as P 1 and P 2 , and he then denoted the progeny resulting from the crossing as the filial, or F 1 , generation. Although the plants of the F 1 generation looked like one parent of the P generation, they were actually hybrids of two different parent plants. Upon observing the uniformity of the F 1 generation, Mendel wondered whether the F 1 generation could still possess the nondominant traits of the other parent in some hidden way.
To understand whether traits were hidden in the F 1 generation, Mendel returned to the method of self-fertilization. Here, he created an F 2 generation by letting an F 1 pea plant self-fertilize (F 1 x F 1 ). This way, he knew he was crossing two plants of the exact same genotype . This technique, which involves looking at a single trait, is today called a monohybrid cross . The resulting F 2 generation had seeds that were either round or wrinkled. Figure 4 shows an example of Mendel's data.
When looking at the figure, notice that for each F 1 plant, the self-fertilization resulted in more round than wrinkled seeds among the F 2 progeny. These results illustrate several important aspects of scientific data:
- Multiple trials are necessary to see patterns in experimental data.
- There is a lot of variation in the measurements of one experiment.
- A large sample size, or "N," is required to make any quantitative comparisons or conclusions.
In Figure 4, the result of Experiment 1 shows that the single characteristic of seed shape was expressed in two different forms in the F 2 generation: either round or wrinkled. Also, when Mendel averaged the relative proportion of round and wrinkled seeds across all F 2 progeny sets, he found that round was consistently three times more frequent than wrinkled. This 3:1 proportion resulting from F 1 x F 1 crosses suggested there was a hidden recessive form of the trait. Mendel recognized that this recessive trait was carried down to the F 2 generation from the earlier P generation .
Mendel and Alleles
As mentioned, Mendel's data did not support the ideas about trait blending that were popular among the biologists of his time. As there were never any semi-wrinkled seeds or greenish-yellow seeds, for example, in the F 2 generation, Mendel concluded that blending should not be the expected outcome of parental trait combinations. Mendel instead hypothesized that each parent contributes some particulate matter to the offspring. He called this heritable substance "elementen." (Remember, in 1865, Mendel did not know about DNA or genes.) Indeed, for each of the traits he examined, Mendel focused on how the elementen that determined that trait was distributed among progeny. We now know that a single gene controls seed form, while another controls color, and so on, and that elementen is actually the assembly of physical genes located on chromosomes. Multiple forms of those genes, known as alleles , represent the different traits. For example, one allele results in round seeds, and another allele specifies wrinkled seeds.
One of the most impressive things about Mendel's thinking lies in the notation that he used to represent his data. Mendel's notation of a capital and a lowercase letter ( Aa ) for the hybrid genotype actually represented what we now know as the two alleles of one gene : A and a . Moreover, as previously mentioned, in all cases, Mendel saw approximately a 3:1 ratio of one phenotype to another. When one parent carried all the dominant traits ( AA ), the F 1 hybrids were "indistinguishable" from that parent. However, even though these F 1 plants had the same phenotype as the dominant P 1 parents, they possessed a hybrid genotype ( Aa ) that carried the potential to look like the recessive P 1 parent ( aa ). After observing this potential to express a trait without showing the phenotype, Mendel put forth his second principle of inheritance: the principle of segregation . According to this principle, the "particles" (or alleles as we now know them) that determine traits are separated into gametes during meiosis , and meiosis produces equal numbers of egg or sperm cells that contain each allele (Figure 5).
Dihybrid Crosses
Mendel had thus determined what happens when two plants that are hybrid for one trait are crossed with each other, but he also wanted to determine what happens when two plants that are each hybrid for two traits are crossed. Mendel therefore decided to examine the inheritance of two characteristics at once. Based on the concept of segregation , he predicted that traits must sort into gametes separately. By extrapolating from his earlier data, Mendel also predicted that the inheritance of one characteristic did not affect the inheritance of a different characteristic.
Mendel tested this idea of trait independence with more complex crosses. First, he generated plants that were purebred for two characteristics, such as seed color (yellow and green) and seed shape (round and wrinkled). These plants would serve as the P 1 generation for the experiment. In this case, Mendel crossed the plants with wrinkled and yellow seeds ( rrYY ) with plants with round, green seeds ( RRyy ). From his earlier monohybrid crosses, Mendel knew which traits were dominant: round and yellow. So, in the F 1 generation, he expected all round, yellow seeds from crossing these purebred varieties, and that is exactly what he observed. Mendel knew that each of the F 1 progeny were dihybrids; in other words, they contained both alleles for each characteristic ( RrYy ). He then crossed individual F 1 plants (with genotypes RrYy ) with one another. This is called a dihybrid cross . Mendel's results from this cross were as follows:
- 315 plants with round, yellow seeds
- 108 plants with round, green seeds
- 101 plants with wrinkled, yellow seeds
- 32 plants with wrinkled, green seeds
Thus, the various phenotypes were present in a 9:3:3:1 ratio (Figure 6).
Next, Mendel went through his data and examined each characteristic separately. He compared the total numbers of round versus wrinkled and yellow versus green peas, as shown in Tables 1 and 2.
Table 1: Data Regarding Seed Shape
Table 2: Data Regarding Pea Color
The proportion of each trait was still approximately 3:1 for both seed shape and seed color. In other words, the resulting seed shape and seed color looked as if they had come from two parallel monohybrid crosses; even though two characteristics were involved in one cross, these traits behaved as though they had segregated independently. From these data, Mendel developed the third principle of inheritance: the principle of independent assortment . According to this principle, alleles at one locus segregate into gametes independently of alleles at other loci. Such gametes are formed in equal frequencies.
Mendel’s Legacy
More lasting than the pea data Mendel presented in 1862 has been his methodical hypothesis testing and careful application of mathematical models to the study of biological inheritance. From his first experiments with monohybrid crosses, Mendel formed statistical predictions about trait inheritance that he could test with more complex experiments of dihybrid and even trihybrid crosses. This method of developing statistical expectations about inheritance data is one of the most significant contributions Mendel made to biology.
But do all organisms pass their on genes in the same way as the garden pea plant? The answer to that question is no, but many organisms do indeed show inheritance patterns similar to the seminal ones described by Mendel in the pea. In fact, the three principles of inheritance that Mendel laid out have had far greater impact than his original data from pea plant manipulations. To this day, scientists use Mendel's principles to explain the most basic phenomena of inheritance.
References and Recommended Reading
- Add Content to Group
Article History
Flag inappropriate.


Email your Friend

- | Lead Editor: Terry McGuire

Within this Subject (29)
- Gene Linkage (5)
- Methods for Studying Inheritance Patterns (7)
- The Foundation of Inheritance Studies (11)
- Variation in Gene Expression (6)
Other Topic Rooms
- Gene Inheritance and Transmission
- Gene Expression and Regulation
- Nucleic Acid Structure and Function
- Chromosomes and Cytogenetics
- Evolutionary Genetics
- Population and Quantitative Genetics
- Genes and Disease
- Genetics and Society
- Cell Origins and Metabolism
- Proteins and Gene Expression
- Subcellular Compartments
- Cell Communication
- Cell Cycle and Cell Division
© 2014 Nature Education
- Press Room |
- Terms of Use |
- Privacy Notice |

Visual Browse

- Introduction to Ecology; Major patterns in Earth’s climate
- Behavioral Ecology
- Population Growth Models
- Survival and Reproduction at the Population Level
- Resources and Competition
- Predation and Community Diversity
- Food webs and Primary Production
- Energy Flow and Nutrient Cycles
- Strong Inference
- What is life?
- What is evolution?
- Evolution by Natural Selection
- Other Mechanisms of Evolution
- Population Genetics: the Hardy-Weinberg Principle
- Phylogenetic Trees
- Earth History and History of Life on Earth
- Origin of Life on Earth
- Gene expression: DNA to protein
- Gene regulation
- Cell division: mitosis and meiosis
Mendelian Genetics
- Chromosome theory of inheritance
- Patterns of inheritance
- Chemical context for biology: origin of life and chemical evolution
- Biological molecules
- Membranes and Transport
- Energy and enzymes
- Respiration, chemiosmosis and oxidative phosphorylation
- Oxidative pathways: electrons from food to electron carriers
- Fermentation, mitochondria and regulation
- Why are plants green, and how did chlorophyll take over the world? (Converting light energy into chemical energy)
- Carbon fixation
- Recombinant DNA
- Cloning and Stem Cells
- Adaptive Immunity
- Human evolution and adaptation
Learning Objectives
- Know and use the vocabulary needed to discuss genetic inheritance including gene, allele, dominant, recessive, gamete, genotype, phenotype, homozygote, heterozygote, carrier
- Explain how chromosomal separation at meiosis leads to segregation of alleles in gametes
- Explain how alignment at metaphase results in independent assortment of (unlinked) genes
- Construct and use a Punnett square for a single trait and for two traits using appropriate terminology
- Determine possible offspring types and phenotypic ratios using probability rules
The terminology of Mendelian inheritance
Gregor Mendel is famous for discovering “particulate inheritance” or the idea that hereditary elements are passed on in discrete units rather than “blended” together at each new generation. Today we call those discrete units genes .
- A gene is a hereditary factor that determines (or influences) a particular trait. A gene is comprised of a specific DNA sequence and is located on a specific region of a specific chromosome. Because of its specific location, a gene can also be called a genetic locus.
- An allele is a particular variant of a gene, in the same way that chocolate and vanilla are particular variants of ice cream.
- An organism’s genotype is the particular collection of alleles found in its DNA. An organism with two of the same alleles for a particular gene is homozygous at that locus; an organism with two different alleles for a particular gene is heterozygous at that locus.
- An organism’s phenotype is its observable traits. An organism can have a heterozygous at a particular locus but have a phenotype that looks like only one of the two alleles. This is because some alleles mask the appearance of others in a dominant/recessive pattern.
- A dominant allele produces its phenotype whether the organism is homozygous or heterozygous at that locus. For example, in humans the allele for brown eyes is dominant to the allele for blue eyes, so a person who is heterozygous at the eye color locus will have brown eyes.
- A recessive allele produces its phenotype only when homozygous at the locus; its phenotype is masked if the locus is heterozygous. For example, a person must have two copies of the blue eye color allele to have blue eyes.
- Sometimes specific recessive alleles are associated with diseases or conditions . A person who is heterozygous for the gene will be phenotypically normal but carry a copy of the recessive, disease-associated allele. This person is said to be a carrier and can pass on the disease allele to his or her offspring.
Crosses with a single trait & the principle of segregation
All of the concepts above are illustrated in the types of experiments that Mendel carried out with pea plants. Pea plants aren’t a particularly exciting organism to study, but they were very useful in figuring out basic patterns of inheritance! The reason they were so useful is that they have a lot of traits that are caused by a single gene with a simple dominant/recessive inheritance pattern (this is actually pretty rare in general – but more on that later ). So what does that statement in bold mean? A classic example is pea shape. Peas can be either round or wrinkly, but not anything in between. Whether they are round or wrinkly is controlled by a single gene with two alleles, and the round allele is dominant to the wrinkly allele. The inheritance pattern if you cross homozygous round and homozygous wrinkly pea plants is illustrated here:

You can see in the first generation (F1) that all offspring produce round seeds, even though they both the round and wrinkly alleles. If the F1 generation self-fertilizes (pea plants – like most plants – produce both male and female gametes), then you now see some offspring that produce rounds seeds and some that produce wrinkly seeds. The round:wrinkly seed producers exist in approximately a 3:1 ratio, illustrated by constructing a Punnett square.
Punnett squares illustrate the fact that each pea plant gamete contains only one allele for each trait. Even though adult pea plants have two copies of each allele, these two alleles become into different gametes. Thus when two gametes come together to create a new plant, each gamete carries one allele resulting in two alleles in the new plant. The idea that each gamete carries only one allele for each trait is the principle of segregation ; that is, the two alleles for a particular trait are segregated into different gametes.
Crosses with two traits and the principle of independent assortment
Pea plants have a lot of other traits beyond seed shape, and Mendel studied seven other traits. Things become more complex when you follow more than one trait at at time . Here is a cross looking at both pea shape (round or wrinkly) and pea color (yellow or green). Follow the logic of the cross below to see why offspring demonstrate a 9:3:3:1 ratio of different phenotypes.

Punnett squares that show two or more traits illustrate the idea that alleles for different traits (different genes) are segregated independently of each other. Yellow seeds are not always round, and green seeds are not always wrinkly; there can be yellow wrinkly seeds, yellow round seeds, green wrinkly seeds, and green round seeds. The idea that alleles for different traits are segregated independently is the principle of independent assortment .
Mendel’s laws and meiosis
Mendel’s laws (principles) of segregation and independent assortment are both explained by the physical behavior of chromosomes during meiosis.
Segregation occurs because each gamete inherits only one copy of each chromosome. Each chromosome has only one copy of each gene; therefore each gamete only gets one allele. Segregation occurs when the homologous chromosomes separate during meiotic anaphase I . This principle is illustrated here:

Independent assortment occurs because homologous chromosomes are randomly segregated into different gametes; ie, one gamete does not only get all maternal chromosomes while the other gets all paternal chromosomes. Independent assortment occurs when homologous chromosomes align randomly at the metaphase plate during meiotic metaphase I . This principle is illustrated here:

Random, independent assortment during metaphase I can be demonstrated by considering a cell with a set of two chromosomes ( n = 2). In this case, there are two possible arrangements at the equatorial plane in metaphase I. The total possible number of different gametes is 2^ n , where n equals the number of chromosomes in a set. In this example, there are four possible genetic combinations for the gametes. With n = 23 in human cells, there are over 8 million possible combinations of paternal and maternal genotypes in a potential offspring.
In class, we will use the information discussed above to determine possible offspring types and phenotypic ratios using simple probability rules. For crosses that involve 2 or more independently assorting traits, using probability rules can be much faster and easier than using 4 x 4 Punnett squares (for 2-factor crosses) or 8 x 8 Punnett squares (for 3-factor crosses. The number of possible gametes is 2^N, where N is the number of factors (genes), and the size of the Punnett square needed is 2^N x 2^N! So instead, we can calculate the outcomes for each factor or gene, then multiply the results.
Example: a cross of AaBbCcDd x AaBbCcDd, where A, B, C and D are 4 different genes, with the dominant alleles given as A, B, C, and D, and the recessive alleles are a, b, c, and d, respectively. What proportion of the progeny will have the dominant phenotype for A and B, and recessive for c and d?
If we look at just Aa x Aa, we know that 3/4 of the progeny will have the dominant A phenotype.
Similarly, for just Bb x Bb, 3/4 of the progeny will have the dominant B phenotype. For Cc x Cc, 1/4 of the progeny will have the recessive c phenotype (cc genotype). For Dd x Dd, 1/4 of the progeny will have the recessive d phenotype (dd genotype). The rules of probability say that, if these genes are sorting independently, we can just multiply these proportions:
The proportion of ABcd phenotype among the progeny = 3/4 x 3/4 x 1/4 x 1/4 = 9/256
Here’s a quick summary of many of these ideas from Ted Ed:
and here is Khan Academy’s take:

UN Sustainable Development Goal (SDG) 3: Good Health and Wellbeing- Understanding genetic inheritance is important for identifying, diagnosing, and treating genetic disorders.
Leave a Reply Cancel reply
You must be logged in to post a comment.
- Entries RSS
- Comments RSS
- Sites@GeorgiaTech
- Search for:
Creative Commons License

- eradication of Asian tiger mosquitos
- gut bacteria and stunted growth
- gut pathogens and autoimmune disease
- human milk oligosaccharides
- Legionella and mitochondria
- mannosides and uropathogenic E. coli
- origin of sickle cell mutation
- trehalose and C. difficile
- Uncategorized

Want to create or adapt books like this? Learn more about how Pressbooks supports open publishing practices.
Chapter 18. Mendelian Genetics

Chapter Outline
- 18.1 Mendel’s Experiments
- 18.2 Mendel’s Principles of Inheritance
- 18.3 Exceptions to Mendel’s Principles of Inheritance
Introduction

Genetics is the study of heredity. Johann Gregor Mendel (1822–1884) set the framework for genetics long before chromosomes or genes had been identified, at a time when meiosis was not well understood ( Figure 18.2 ). Mendel selected a simple biological system and conducted methodical, quantitative analyses using large sample sizes. Because of Mendel’s work, the fundamental principles of heredity were revealed. We now know that genes, carried on chromosomes, are the basic functional units of heredity with the capability to be replicated, expressed, or mutated. Today, the postulates put forth by Mendel form the basis of classical, or Mendelian, genetics. Not all genes are transmitted from parents to offspring according to Mendelian genetics, but Mendel’s experiments serve as an excellent starting point for thinking about inheritance.
18.1 | Mendel’s Experiments
Learning Objectives
By the end of this section, you will be able to:
- Describe the scientific reasons for the success of Mendel’s experimental work.
- Describe the expected outcomes of monohybrid crosses involving dominant and recessive alleles.
Johann Gregor Mendel (1822–1884) was a lifelong learner, teacher, scientist, and man of faith. As a young adult, he joined the Augustinian Abbey of St. Thomas in Brno in what is now the Czech Republic. Supported by the monastery, he taught physics, botany, and natural science courses at the secondary and university levels. In 1856, he began a decade-long research pursuit involving inheritance patterns in honeybees and plants, ultimately settling on pea plants as his primary model system . In 1865, Mendel presented the results of his experiments with nearly 30,000 pea plants to the local Natural History Society. He demonstrated that traits are transmitted faithfully from parents to offspring independently of other traits and in dominant and recessive patterns. In 1866, he published his work, Exp eriments in Plant Hybridization [1] in the proceedings of the Natural History Society of Brünn.
Mendel’s work went virtually unnoticed by the scientific community that believed, incorrectly, that the process of inheritance involved a blending of parental traits that produced an intermediate physical appearance in offspring; this hypothetical process appeared to be correct because of what we know now as continuous variation. Continuous variation results when many genes work together to determine a characteristic, such as human height or eye color. Offspring appear to be a “blend” of their parents’ traits when we look at characteristics that exhibit continuous variation.
Mendel worked with traits that were inherited in distinct classes, such as violet versus white flowers. These traits display discontinuous variation . Mendel’s choice of these kinds of traits allowed him to see that the traits were not blended in the offspring, nor were they absorbed, but rather that they kept their distinctness and could be passed on. In 1868, Mendel became abbot of the monastery and exchanged his scientific pursuits for his pastoral duties. He was not recognized for his extraordinary scientific contributions during his lifetime. In fact, it was not until 1900 that his work was rediscovered, reproduced, and revitalized by scientists on the brink of discovering the chromosomal basis of heredity.
18.1.1 Mendel’s Model System
Mendel’s seminal work was accomplished using the garden pea, Pisum sativum , to study inheritance. This species naturally self-fertilizes, such that pollen encounters ova within individual flowers. The flower petals remain sealed tightly until after pollination, preventing pollination from other plants. The result is highly inbred, or “true-breeding,” pea plants. These are plants that always produce offspring that look like the parent. By experimenting with true-breeding pea plants, Mendel avoided the appearance of unexpected traits in offspring that might occur if the plants were not true breeding. The garden pea also grows to maturity within one season, meaning that several generations could be evaluated over a relatively short time. Finally, large quantities of garden peas could be cultivated simultaneously, allowing Mendel to conclude that his results did not come about simply by chance.
18.1.2 Mendelian Crosses
Mendel performed hybridizations , which involve mating two true-breeding individuals that have different traits. In the pea, this is done by manually transferring pollen from one pea plant to the stigma of another pea plant. In plants, pollen carries the male gametes (sperm) to the stigma, a sticky organ that traps pollen and allows the sperm to move down the pistil to the female gametes (ova) below. To prevent the pea plant that was receiving pollen from self-fertilizing and confounding his results, Mendel painstakingly removed all of the pollen-producing anthers from the plant’s flowers before they had a chance to mature.
Plants used in first-generation crosses were called P , or parental generation, plants ( Figure 18.3 ). Mendel collected the seeds that resulted from each cross and grew them the following season. These offspring were called the F1 , or the first filial ( filial = offspring, daughter or son), generation. Once Mendel examined the characteristics in the F1 generation of plants, he allowed them to self-fertilize. He then collected and grew the seeds from the F1 plants to produce the F2 , or second filial, generation. Mendel’s experiments extended beyond the F2 generation to the F3 and F4 generations, and so on, but it was the ratio of characteristics in the P−F1−F2 generations that were the most intriguing and became the basis for Mendel’s principles.

18.1.3 Garden Pea Characteristics Revealed the Basics of Heredity
In his 1865 publication, Mendel reported the results of his crosses involving seven different characteristics, each with two contrasting traits. A trait is defined as a variation in the physical appearance of a heritable characteristic. The characteristics include: tall vs. short plant height, wrinkled vs. round seeds, green vs. yellow seeds, violet vs. white flowers, etc. ( Table 18.1 ). To fully examine each characteristic, Mendel generated large numbers of F1 and F2 plants, reporting results from 19,959 F2 plants alone.
As an example, let us look at Mendel’s results for the flower color trait. First, Mendel confirmed that he had plants that bred true for white or violet flower color. Regardless of how many generations Mendel examined, all self-crossed offspring of parents with white flowers had white flowers, and all self-crossed offspring of parents with violet flowers had violet flowers. In addition, Mendel confirmed that, other than flower color, the pea plants were physically identical.
Once these validations were complete, Mendel applied pollen from a plant with violet flowers to the stigma of a plant with white flowers. After gathering and sowing the seeds that resulted from this cross, Mendel found that 100 percent of the F1 hybrid generation had violet flowers. Conventional wisdom at that time would have predicted the hybrid flowers to be pale violet or for hybrid plants to have equal numbers of white and violet flowers. In other words, the contrasting parental traits were expected to blend in the offspring. Instead, Mendel’s results demonstrated that the white flower trait in the F1 generation had completely disappeared.
Importantly, Mendel did not stop his experimentation there. He allowed the F1 plants to self-fertilize and found that, of F2- generation plants, 705 had violet flowers and 224 had white flowers. This was a ratio of 3.15 violet flowers per one white flower, or approximately 3:1. When Mendel transferred pollen from a plant with violet flowers to the stigma of a plant with white flowers and vice versa, he obtained about the same ratio regardless of which parent, male or female, contributed which trait. This is called a reciprocal cross —a paired cross in which the respective traits of the male and female in one cross become the respective traits of the female and male in the other cross. For the other six characteristics Mendel examined, the F1 and F2 generations behaved in the same way as they had for flower color. One of the two traits would disappear completely from the F1 generation only to reappear in the F2 generation at a ratio of approximately 3:1 ( Table 18.1 ).
Table 18.1 The Results of Mendel’s Garden Pea Hybridizations
|
|
|
|
|
|
| Flower color | Violet vs. white | 100 percent violet | 705 violet 224 white | 3.15:1 |
| Flower position | Axial vs. terminal | 100 percent axial | 651 axial 207 terminal | 3.14:1 |
| Plant height | Tall vs. dwarf | 100 percent tall | 787 tall 277 dwarf | 2.84:1 |
| Seed texture | Round vs. wrinkled | 100 percent round | 5,474 round 1,850 wrinkled | 2.96:1 |
| Seed color | Yellow vs. green | 100 percent yellow | 6,022 yellow 2,001 green | 3.01:1 |
| Pea pod texture | Inflated vs. constricted | 100 percent inflated | 882 inflated 299 constricted | 2.95:1 |
| Pea pod color | Green vs. yellow | 100 percent green | 428 green 152 yellow | 2.82:1 |
18.2 | Mendel’s Principles of Inheritance
- Describe the three principles of inheritance.
- Explain the relationship between phenotype and genotype.
- Develop a Punnett square to calculate the expected proportions of genotypes and phenotypes in a monohybrid cross.
- Explain the purpose and methods of a test cross.
- Draw and interpret a pedigree.
Mendel generalized the results of his pea-plant experiments into three principles that describe the basis of inheritance in diploid organisms. They are: the principle of segregation, the principle of dominance, and the principle of independent assortment. Together, these principles summarize the basics of classical, or Mendelian, genetics.
18.2.1 The Principle of Segregation
Since the white flower trait reappeared in the F2 generation, Mendel saw that the traits remained separate (not blended) in the plants of the F1 generation. This led to the principle of segregation , which states that individuals have two copies of each trait, and that each parent transmits one of its two copies to its offspring.
We now know that the traits that are passed on are a result of genes that are inherited on chromosomes during meiosis and fertilization. The fact that the genetic factors proposed by Mendel were carried on chromosomes was proposed in 1902 by Walter and Sutton and Theodor Boveri ( Figure 18.4 ) as the Chromosomal Theory of Inheritance .

Different versions of genes are called alleles . Diploid organisms that have two identical alleles of a gene on their two homologous chromosomes are homozygous for that trait. Diploid organisms that have two different alleles of a gene on their two homologous chromosomes are heterozygous for that trait.
The physical basis of the principle of segregation is the first division of meiosis, in which the homologous chromosomes with their different versions of each gene are segregated into daughter nuclei. Since each gamete receives only one homolog of each chromosome, it follows that they receive only one allele for each trait. At fertilization, the zygote receives one of each homologous chromosome, and one of each allele, from each parent.
18.2.2 The Principle of Dominance
Upon compiling his results for many thousands of plants, Mendel concluded that the characteristics could be divided into dominant and recessive traits. Dominant traits are those that are expressed in a hybridization. Recessive traits become latent, or disappear, in the offspring of a hybridization but reappear in the progeny of the hybrid offspring. Thus, the violet-flower trait is dominant and the white-flower trait is recessive.

The principle of dominance states that in a heterozygote, only the dominant allele will be expressed. The recessive allele will remain “latent” but will be transmitted to offspring by the same manner in which the dominant allele is transmitted. The recessive trait will only be expressed by offspring that have two copies of this allele ( Figure 18. 5 ). Individuals with a dominant trait could have either two dominant versions of the trait or one dominant and one recessive version of the trait. Individuals with a recessive trait have two recessive alleles.
In Mendel’s experiments, the principle of dominance explains why the F1 heterozygous offspring were identical to one of the parents, rather than expressing both alleles. For a gene that is expressed in a dominant and recessive pattern, homozygous dominant and heterozygous organisms will look identical. The recessive allele will only be observed in homozygous recessive individuals. Some examples of human dominant and recessive traits are shown in Table 18.2 .
Table 18.2 Examples of dominant and recessive traits in humans.
|
| |
| Achondroplasia | Albinism |
| Brachydactyly | Cystic fibrosis |
| Huntington’s disease | Duchenne muscular dystrophy |
| Marfan syndrome | Galactosemia |
| Neurofibromatosis | Phenylketonuria |
| Widow’s peak | Sickle-cell anemia |
| Wooly hair | Tay-Sachs disease |
The principles of segregation and dominance could be deduced by simple crosses that follow only one genetic trait. These crosses are called monohybrid crosses . Before we discuss the principle of independent assortment, let’s look at some tools and terminology used for monohybrid crosses.
18.2.3 Phenotypes and Genotypes
Several conventions exist for referring to genes and alleles. For the purposes of this chapter, we will abbreviate genes using the first letter of the gene’s corresponding dominant trait. For example, green is the dominant trait for pea pod color, so the pod-color gene would be abbreviated as G (note that it is customary to italicize gene designations). Furthermore, we will use uppercase and lowercase letters to represent dominant and recessive alleles, respectively. Therefore, we would refer to the genotype of a homozygous dominant pea plant with green pods as GG , a homozygous recessive pea plant with yellow pods as gg , and a heterozygous pea plant with green pods as Gg .
The two alleles for each given gene in a diploid organism may be expressed and interact to produce physical characteristics. The observable traits expressed by an organism are referred to as its phenotype . An organism’s underlying genetic makeup, which alleles it has, is called its genotype . Mendel’s hybridization experiments demonstrate the difference between phenotype and genotype. When true-breeding plants in which one parent had yellow pods and one had green pods were cross-fertilized, all of the F1 hybrid offspring had green pods. Although the hybrid offspring had the same phenotype as the true-breeding parent with green pods, we know that the genotype of the parent was homozygous dominant ( GG ), while the genotype of the F1 offspring was heterozygous ( Gg ). We know this since the yellow pod allele reappeared in some of the F2 offspring ( gg ).
18.2.4 Using Punnett Squares for Monohybrid Crosses
Punnett squares , devised by the British geneticist Reginald Punnett, can be used to predict the possible outcomes of a genetic cross or mating and their expected frequencies. To demonstrate a monohybrid cross, consider the case of true- breeding pea plants with yellow versus green pea seeds. The dominant seed color is yellow; therefore, the parental genotypes were YY for the plants with yellow seeds and yy for the plants with green seeds, respectively. To prepare a Punnett square, all possible combinations of the parental alleles are listed along the top (for one parent) and side (for the other parent) of a grid, representing their meiotic segregation into haploid gametes. Then the combinations of egg and sperm are made in the boxes in the table to show which alleles are combining. Each box then represents the diploid genotype of a zygote, or fertilized egg, that could result from this mating. Because each possibility is equally likely, genotypic ratios can be determined from a Punnett square. If the pattern of inheritance (dominant or recessive) is known, the phenotypic ratios can be inferred as well. For a monohybrid cross of two true-breeding parents, each parent contributes one type of allele. In this case, only one genotype is possible. All offspring are Yy and have yellow seeds ( Figure 18. 6 ).

A self-cross of one of the Yy heterozygous offspring can be represented in a 2 × 2 Punnett square because each parent can donate one of two different alleles. Therefore, the offspring can potentially have one of four allele combinations: YY , Yy , yY , or yy ( Figure 18. 6 ). Notice that there are two ways to obtain the Yy genotype: a Y from the egg and a y from the sperm, or a y from the egg and a Y from the sperm. Both of these possibilities must be counted. Recall that Mendel’s pea- plant characteristics behaved in the same way in reciprocal crosses. Therefore, the two possible heterozygous combinations produce offspring that are genotypically and phenotypically identical despite their dominant and recessive alleles deriving from different parents.
Because fertilization is a random event, we expect each combination to be equally likely and for the offspring to exhibit a ratio of YY : Yy : yy genotypes of 1:2:1 ( Figure 18. 6 ). Furthermore, because the YY and Yy offspring have yellow seeds and are phenotypically identical, we expect the offspring to exhibit a phenotypic ratio of 3 yellow:1 green. Indeed, working with large sample sizes, Mendel observed approximately this ratio in every F2 generation resulting from crosses for individual traits.
Using a Test Cross to Determine Genotype
Beyond predicting the offspring of a cross between known homozygous or heterozygous parents, Mendel also developed a way to determine whether an organism that expressed a dominant trait was a heterozygote or a homozygote. Called the test cross , this technique is still used by plant and animal breeders. In a test cross, an organism with the dominant phenotype is crossed with an organism that is homozygous recessive for the same characteristic. If the dominant- expressing organism is a homozygote, then all F1 offspring will be heterozygotes expressing the dominant trait. Alternatively, if the dominant expressing organism is a heterozygote, the F1 offspring will exhibit a 1:1 ratio of heterozygotes and recessive homozygotes ( Figure 18. 7 ). The test cross further validates Mendel’s postulate that pairs of unit factors segregate equally.

Concept Check
In pea plants, round peas (R) are dominant to wrinkled peas (r). You do a test cross between a pea plant with wrinkled peas (genotype rr) and a plant of unknown genotype that has round peas. You end up with three plants, all which have round peas.
- From this data, can you tell if the round pea parent plant is homozygous dominant or heterozygous?
- If the round pea parent plant is heterozygous, what is the probability that a random sample of 3 progeny peas will all be round?
18.2.5 Using Pedigrees to Study Inheritance Patterns
Many human diseases are inherited genetically. A healthy person in a family in which some members suffer from a recessive genetic disorder may want to know if he or she has the disease-causing gene and what risk exists of passing the disorder on to his or her offspring. Of course, doing a test cross in humans is unethical and impractical. Instead, geneticists use pedigree analysis to study the inheritance pattern of human genetic diseases.
Each row of a pedigree represents one generation of the family. Women are represented by circles; males by squares. People who had children together are connected with a horizontal line and their children are connected to this line with a vertical line. See Figure 18. 8 for an example of a pedigree for a human genetic disease.
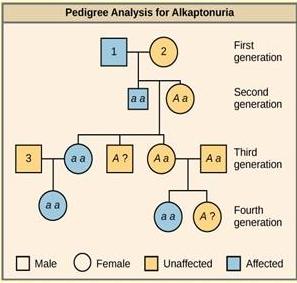
People with the recessive genetic disease alkaptonuria cannot properly metabolize two amino acids, phenylalanine and tyrosine. Affected individuals may have darkened skin and brown urine, and may suffer joint damage and other complications.
In this pedigree, individuals with the disorder are indicated in blue and have the genotype aa . Unaffected individuals are indicated in yellow and have the genotype AA or Aa . Note that it is often possible to determine a person’s genotype from the genotype of their offspring. For example, if neither parent has the disorder but their child does, both parents must be heterozygous. Two individuals on the pedigree have an unaffected phenotype but unknown genotype. Because they do not have the disorder, they must have at least one normal allele, so their genotype gets the “ A? ” designation.
What are the genotypes of the individuals labeled 1, 2, and 3?
18.2.6 Principle of Independent Assortment
Mendel’s principle of independent assortment states that genes do not influence each other with regard to the sorting of alleles into gametes, and every possible combination of alleles for every gene is equally likely to occur. The independent assortment of genes can be illustrated by a dihybrid cross, a cross between two true-breeding parents that express different traits for two characteristics. Consider the characteristics of seed color and seed texture for two pea plants, one that has green, wrinkled seeds ( yyrr ) and another that has yellow, round seeds ( YYRR ). Because each parent is homozygous, the principle of segregation indicates that the gametes for the green/wrinkled plant all are yr , and the gametes for the yellow/round plant are all YR . Therefore, the F1 generation of offspring all are YyRr ( Figure 18.9 ).
For the F2 generation, the principle of segregation requires that each gamete receive either an R allele or an r allele along with either a Y allele or a y allele. The principle of independent assortment states that a gamete into which an r allele sorted would be equally likely to contain either a Y allele or a y allele. Thus, there are four equally likely gametes that can be formed when the YyRr heterozygote is self-crossed, as follows: YR , Yr , yR , and yr . Arranging these gametes along the top and left of a 4 × 4 Punnett square gives us 16 equally likely genotypic combinations. From these genotypes, we infer a phenotypic ratio of 9 round/yellow:3 round/green:3 wrinkled/yellow:1 wrinkled/green ( Figure 18.9 ).
The physical basis for the principle of independent assortment also lies in meiosis I, in which the different homologous pairs line up in random orientations. Each gamete can contain any combination of paternal and maternal chromosomes (and therefore the genes on them) because the orientation of tetrads on the metaphase plane is random.

Testing the Hypothesis of Independent Assortment
To better appreciate the amount of labor and ingenuity that went into Mendel’s experiments, proceed through one of Mendel’s dihybrid crosses.
Question : What will be the offspring of a dihybrid cross?
Background : Consider that you have access to a large garden in which you can cultivate thousands of pea plants. There are several true-breeding plants with the following pairs of traits: tall plants with inflated pods, and dwarf plants with constricted pods. Before the plants have matured, you remove the pollen-producing organs from the tall/inflated plants in your crosses to prevent self-fertilization. When the plants mature, they are manually crossed by transferring pollen from the dwarf/constricted plants to the stigmata of the tall/inflated plants.
Hypothesis : Both trait pairs will sort independently according to Mendelian principles. When the true-breeding parents are crossed, all of the F1 offspring are tall and have inflated pods, which indicates that the tall (T ) and inflated (I) traits are dominant over the dwarf (t) and constricted (i) traits, respectively. A self-cross of the F1 heterozygotes results in 2,000 F2 progeny.
Test the hypothesis : You cross the dwarf and tall plants and then self-cross the offspring. For best results, this is repeated with hundreds or even thousands of pea plants. What special precautions should be taken in the crosses and in growing the plants?
If these traits sort independently, the ratios of tall:dwarf and inflated:constricted will each be 3:1. Each member of the F1 generation therefore has a genotype of TtIi . Figure 18.1 0 shows a cross between two TtIi individuals. There are 16 possible offspring genotypes. The offspring proportions: tall/inflated:tall/constricted:dwarf/inflated:dwarf/constricted show a 9:3:3:1 ratio. Notice from the grid that when considering the tall/dwarf and inflated/constricted trait pairs in isolation, they are each inherited in 3:1 ratios.
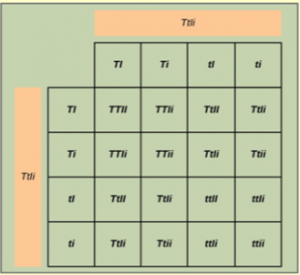
Analyze your data: You observe the following plant phenotypes in the F2 generation: 2706 tall/inflated, 930 tall/constricted, 888 dwarf/inflated, and 300 dwarf/constricted. Reduce these findings to a ratio and determine if they are consistent with Mendelian principles.
Form a conclusion: Were the results close to the expected 9:3:3:1 phenotypic ratio? Do the results support the prediction? What might be observed if far fewer plants were used, given that alleles segregate randomly into gametes? Try to imagine growing that many pea plants, and consider the potential for experimental error. For instance, what would happen if it was extremely windy one day?
18.3 | Exceptions to Mendel’s Principles of Inheritance
- Identify non-Mendelian inheritance patterns such as incomplete dominance, codominance, and sex linkage.
- Describe genetic linkage.
- Describe how chromosome maps are created.
- Explain the phenotypic outcomes of epistatic effects between genes.
Although Mendel’s principles still apply to some situations, many situations exist in which they do not apply. These “exceptions” to Mendelian genetics are discussed below.
18.3.1 Alternatives to Dominance and Recessiveness
Since Mendel’s experiments with pea plants, other researchers have found that the principle of dominance does not always hold true. Instead, several different patterns of inheritance have been found to exist.
Incomplete Dominance

Mendel’s results, that traits are inherited as dominant and recessive pairs, contradicted the view at that time that offspring exhibited a blend of their parents’ traits. However, the heterozygote phenotype occasionally does appear to be intermediate between the two parents. For example, in the snapdragon, Antirrhinum majus ( Figure 18. 11 ), a cross between a homozygous parent with white flowers ( CWCW ) and a homozygous parent with red flowers ( CRCR ) will produce offspring with pink flowers ( CRCW ). (Note that different genotypic abbreviations are used for Mendelian extensions to distinguish these patterns from simple dominance and recessiveness.) This pattern of inheritance is described as incomplete dominance , denoting the expression of two contrasting alleles such that the individual displays an intermediate phenotype. The allele for red flowers is incompletely dominant over the allele for white flowers. However, the results of a heterozygote self-cross can still be predicted, just as with Mendelian dominant and recessive crosses. In this case, the genotypic ratio would be 1 CRCR :2 CRCW :1 CWCW , and the phenotypic ratio would be 1:2:1 for red:pink:white.
Codominance
A variation on incomplete dominance is codominance , in which both alleles for the same characteristic are simultaneously expressed in the heterozygote. An example of codominance is the MN blood groups of humans. The M and N alleles are expressed in the form of an M or N antigen present on the surface of red blood cells. Homozygotes ( LMLM and LNLN ) express either the M or the N allele, and heterozygotes ( LMLN ) express both alleles equally. In a self-cross between heterozygotes expressing a codominant trait, the three possible offspring genotypes are phenotypically distinct. However, the 1:2:1 genotypic ratio characteristic of a Mendelian monohybrid cross still applies.
Multiple Alleles
Mendel implied that only two alleles, one dominant and one recessive, could exist for a given gene. We now know that this is an oversimplification. Although individual humans (and all diploid organisms) can only have two alleles for a given gene, multiple alleles may exist at the population level such that many combinations of two alleles are observed. Note that when many alleles exist for the same gene, the convention is to denote the most common phenotype or genotype among wild animals as the wild type (often abbreviated “+”); this is considered the standard or norm. All other phenotypes or genotypes are considered variants of this standard, meaning that they deviate from the wild type. The variant may be recessive or dominant to the wild-type allele.

An example of multiple alleles is coat color in rabbits ( Figure 18. 12 ). Here, four alleles exist for the c gene. The wild-type version, C+C+ , is expressed as brown fur. The chinchilla phenotype, cchcch , is expressed as black-tipped white fur. The Himalayan phenotype, chch , has black fur on the extremities and white fur elsewhere. Finally, the albino, or “colorless” phenotype, cc , is expressed as white fur. In cases of multiple alleles, dominance hierarchies can exist. In this case, the wild- type allele is dominant over all the others, chinchilla is incompletely dominant over Himalayan and albino, and Himalayan is dominant over albino. This hierarchy, or allelic series, was revealed by observing the phenotypes of each possible heterozygote offspring.
An example of multiple allelism in humans pertains to ABO blood type. A person’s blood type (e.g., type A or type O) is caused by different combinations of three alleles: IA, IB, and IO. A person with type A blood could have either IAIA or IAIO genotype. A person with type B blood could have IBIB or IBIO genotype. A person with type O blood must have the IOIO genotype. Note that type AB blood is an example of codominance (IAIB).
The complete dominance of a wild-type phenotype over all other mutants often occurs as an effect of “dosage” of a specific gene product, such that the wild-type allele supplies the correct amount of gene product whereas the mutant alleles cannot. For rabbit fur color, the wild-type allele may supply a given dosage of fur pigment, whereas the mutants supply a lesser dosage or none at all.
Multiple Alleles Confer Drug Resistance in the Malaria Parasite
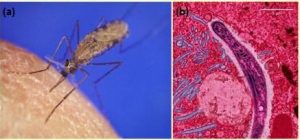
Malaria is a parasitic disease that is transmitted to humans by infected female Anopheles gambiae mosquitos ( Figure 18.13a ). It is characterized by cyclic high fevers, chills, flu-like symptoms, and severe anemia. Plasmodium falciparum is the most deadly causative agent of malaria ( Figure 18.13b ). When promptly and correctly treated, P. falciparum malaria has a mortality rate of 0.1%. However, in some parts of the world, the parasite has evolved resistance to commonly used malaria treatments, so the most effective malarial treatments can vary by geographic region.
In Southeast Asia, Africa, and South America, P. falciparum has developed resistance to the anti-malarial drugs chloroquine, mefloquine, and sulfadoxine-pyrimethamine. P. falciparum , which is haploid during the life stage in which it infects humans, has evolved multiple drug-resistant mutant alleles of the dhps gene. Varying degrees of sulfadoxine resistance are associated with each of these alleles. Being haploid, P. falciparum needs only one drug-resistant allele to express this trait.
Environmental Effects
Interestingly, the Himalayan phenotype in rabbits is the result of an allele that produces a temperature-sensitive gene product that only produces pigment in the cooler extremities of the rabbit’s body. In this case, the protein product of the gene does not fold correctly at high temperatures. A similar gene gives Siamese cats their distinctive coloration.
Temperature-sensitive proteins are also at work in arctic foxes and rabbits, which are white in the winter and darker colored during the summer. In these cases, the protein product of the gene does not fold correctly at colder temperatures. The mutation that caused this coloration was advantageous to these species, so they persisted in the populations.
18.3.2 X-Linked Traits are an Exception to the Principle of Segregation

In humans, as well as in many other animals and some plants, the sex of the individual is determined by sex chromosomes. The sex chromosomes are one pair of non-homologous chromosomes. Until now, we have only considered inheritance patterns among non-sex chromosomes, or autosomes. In addition to 22 homologous pairs of autosomes, human females have a homologous pair of X chromosomes, whereas human males have an XY chromosome pair. Although the Y chromosome contains a small region of similarity to the X chromosome so that they can pair during meiosis, the Y chromosome is much shorter and contains many fewer genes. When a gene is present on the X chromosome, it is said to be X-linked .
Eye color in Drosophila was one of the first X-linked traits to be identified. Like humans, Drosophila males are XY and females are XX. In flies, the wild-type eye color is red (X W ) which is dominant to white eye color (X w ) ( Figure 18.1 4) . Females can be X W X W , X W X w or X w X w . However, Drosophila males lack a second allele copy on the Y chromosome, so their genotype can only be X W Y or X w Y. Males are said to be hemizygous , because they have only one allele for any X- linked characteristic. Hemizygosity makes the descriptions of dominance and recessiveness irrelevant for XY males.
In an X-linked cross, the genotypes of F1 and F2 offspring depend on whether the recessive trait was expressed by the male or the female in the P generation. When the P male expresses the white-eye phenotype and the female is homozygous red-eyed, all members of the F1 generation exhibit red eyes ( Figure 18.1 5 ). The F1 females are heterozygous (X W X w ), and the males are all X W Y, since they received their X chromosome from the homozygous dominant P female and their Y chromosome from the P male. A cross between a X W X w female and an X W Y male would produce only red-eyed females and both red- and white-eyed males. A cross between a homozygous white-eyed female and a male with red eyes would produce only heterozygous red-eyed females and only white-eyed males.
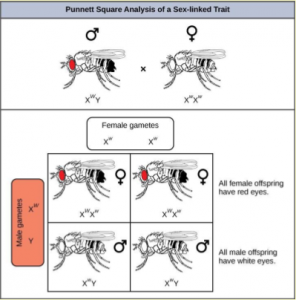
What ratio of offspring would result from a cross between a white-eyed male and a female that is heterozygous for red eye color?
In some groups of organisms with sex chromosomes, the gender with the non-homologous sex chromosomes is the female rather than the male. This is the case for all birds. In this case, sex-linked traits will be more likely to appear in the female, in which they are hemizygous.
Human Sex-linked Disorders
Sex-linkage studies in Morgan’s laboratory provided the fundamentals for understanding X-linked recessive disorders in humans, which included red-green color blindness, Types A and B hemophilia, and muscular dystrophy. Because human males need to inherit only one recessive mutant X allele to be affected, X-linked disorders are disproportionately observed in males. Females must inherit recessive X-linked alleles from both of their parents in order to express the trait. When they inherit one recessive X-linked mutant allele and one dominant X-linked wild-type allele, they are carriers of the trait and are typically unaffected. Carrier females can manifest mild forms of the trait due to the inactivation of the dominant allele located on one of the X chromosomes. However, female carriers can contribute the trait to their sons, resulting in the son exhibiting the trait, or they can contribute the recessive allele to their daughters, resulting in the daughters being carriers of the trait ( Figure 18.1 6 ). Although some Y-linked recessive disorders exist, typically they are associated with infertility in males and are therefore not transmitted to subsequent generations.

18.3.3 Lethal Alleles are Apparent Exceptions to the Principle of Segregation

A large proportion of genes in an individual’s genome are essential for survival. Occasionally, a nonfunctional allele for an essential gene can arise by mutation and be transmitted in a population through heterozygous carriers. The wild-type allele functions at a capacity sufficient to sustain life and is therefore considered to be dominant over the nonfunctional allele. If two heterozygous parents mate, one quarter of their offspring will be homozygous recessive. Because the gene is essential, these individuals will die. This will cause the genotypic ratio among surviving offspring to be 2:1 rather than 3:1. This inheritance pattern is referred to as recessive lethal .
The dominant lethal inheritance pattern is one in which an allele is lethal both in the homozygote and the heterozygote. Dominant lethal alleles are very rare because, as you might expect, the allele only lasts one generation and is not transmitted. However, dominant lethal alleles might not be expressed until adulthood. The allele may be unknowingly passed on, resulting in a delayed death in both generations. An example of this in humans is Huntington disease, in which the nervous system gradually wastes away ( Figure 18.1 7 ). People who are heterozygous for the dominant Huntington allele ( Hh ) will inevitably develop the fatal disease. However, the onset of Huntington disease may not occur until age 40, at which point the afflicted persons may have already passed the allele to 50 percent of their offspring.
18.3.4 Linked Genes Violate the Principle of Independent Assortment
Although all of Mendel’s pea characteristics behaved according to the principle of independent assortment, we now know that some allele combinations are not inherited independently of each other. Genes that are located on different chromosomes will always sort independently. However, each chromosome contains hundreds or thousands of genes, organized linearly on chromosomes like beads on a string. Genes that are on the same chromosome are linked and are therefore likely to be inherited together. When homologs separate during meiosis I, entire chromosomes segregate into separate daughter cells, carrying all of their linked genes with them.
However, because of crossover, it is possible for two genes on the same chromosome to behave independently, or as if they are not linked. To understand this, let’s consider the biological basis of gene linkage and recombination.
Homologous chromosomes possess the same genes in the same order. However, since each homolog came from a different parent, the alleles may differ on homologous chromosome pairs. Prior to meiosis I, homologous chromosomes replicate and synapse so that genes on the homologs align with each other. At this stage, segments of homologous chromosomes cross over and exchange segments of genetic material ( Figure 18.1 8 ). Because the genes are aligned, the gene order is not altered. Instead, the result of recombination is that maternal and paternal alleles are combined onto the same chromosome. Across a given chromosome, several recombination events may occur, causing extensive shuffling of alleles.

When two genes are located in close proximity on the same chromosome, their alleles are more likely to be transmitted through meiosis together. To exemplify this, imagine a dihybrid cross involving flower color and plant height in which the genes are next to each other on the chromosome. If the homologous chromosome from one parent has alleles for tall plants and red flowers, and the homolog from the other parent has alleles for short plants and yellow flowers, then when the gametes are formed, the tall and red alleles will go together into a gamete and the short and yellow alleles will go into other gametes. These are called the parental genotypes because they have been inherited intact from the parents of the individual producing gametes. Since the genes were close together on the same chromosomes, the chance of a crossover event happening between them is slim. Therefore, there will be no gametes with tall and yellow alleles and no gametes with short and red alleles. If you create the Punnett square with these gametes, you will see that the classical Mendelian prediction of a 9:3:3:1 outcome of a dihybrid cross would not apply
As the distance between two genes increases, the probability of crossovers between them increases, and the genes behave more as if they are on separate chromosomes. The further apart two linked genes are on a chromosome, the more progeny with nonparental genotypes will appear.
Genetic Linkage and Distances
Geneticists have used the proportion of nonparental gametes as a measure of how far apart genes are on a chromosome. Using this information, they have constructed elaborate maps of genes on chromosomes. Briefly, the more crossover that occurs between two linked genes, the further apart they are on the chromosome. The frequency of crossover is measured by counting the number of offspring that have nonparental genotypes. By using recombination frequency to predict genetic distance, the relative order of genes on chromosome 2 could be inferred.
18.3.5 Epistasis is an Exception to the Principle of Independent Assortment
Mendel’s studies in pea plants implied that every characteristic was distinctly and completely controlled by a single gene. In fact, single observable characteristics are almost always under the influence of multiple genes (each with two or more alleles) acting in unison. For example, at least eight genes contribute to eye color in humans.
Genes may function in complementary or synergistic fashions, such that two or more genes need to be expressed simultaneously to affect a phenotype. Genes may also oppose each other. In epistasis , the interaction between genes is antagonistic, such that one gene masks or interferes with the expression of another. Often the biochemical basis of epistasis is a gene pathway in which the expression of one gene is dependent on the function of a gene that precedes or follows it in the pathway.
An example of epistasis is pigmentation in mice. The wild-type coat color, agouti ( AA ), is dominant to solid-colored fur ( aa ). However, a separate gene ( C ) is necessary for pigment production. A mouse with a recessive c allele at this locus is unable to produce pigment and is albino regardless of the allele present at locus A . Therefore, the genotypes AAcc , Aacc , and aacc all produce an albino phenotype. A cross between heterozygotes for both genes ( AaCc x AaCc ) would generate offspring with a phenotypic ratio of 9 agouti:3 solid color:4 albino ( Figure 18.19 ). In this case, the C gene is epistatic to the A gene.

Epistasis can also occur when a dominant allele masks expression at a separate gene. Fruit color in summer squash is expressed in this way. Homozygous recessive expression of the W gene ( ww ) coupled with homozygous dominant or heterozygous expression of the Y gene ( YY or Yy ) generates yellow fruit, and the wwyy genotype produces green fruit. However, if a dominant copy of the W gene is present in the homozygous or heterozygous form, the summer squash will produce white fruit regardless of the Y alleles. A cross between white heterozygotes for both genes ( WwYy × WwYy ) would produce offspring with a phenotypic ratio of 12 white:3 yellow:1 green.
Finally, epistasis can be reciprocal such that either gene, when present in the dominant (or recessive) form, expresses the same phenotype. In the shepherd’s purse plant ( Capsella bursa-pastoris ), the characteristic of seed shape is controlled by two genes in a dominant epistatic relationship. When the genes A and B are both homozygous recessive ( aabb ), the seeds are ovoid. If the dominant allele for either of these genes is present, the result is triangular seeds. That is, every possible genotype other than aabb results in triangular seeds, and a cross between heterozygotes for both genes ( AaBb x AaBb ) would yield offspring with a phenotypic ratio of 15 triangular:1 ovoid.
As you work through genetics problems, keep in mind that any single characteristic that results in a phenotypic ratio that totals 16 is typical of a two-gene interaction. Recall the phenotypic inheritance pattern for Mendel’s dihybrid cross, which considered two non-interacting genes—9:3:3:1. Similarly, we would expect interacting gene pairs to also exhibit ratios expressed as 16 parts. Note that we are assuming the interacting genes are not linked; they are still assorting independently into gametes.
- Johann Gregor Mendel, Versuche über Pflanzenhybriden Verhandlungen des naturforschenden Vereines in Brünn, Bd. IV für das Jahr, 1865 Abhandlungen, 3–47. [for English translation see http://www.mendelweb.org/Mendel.plain.html] ↵
Introduction to Molecular and Cell Biology Copyright © 2020 by Katherine R. Mattaini is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License , except where otherwise noted.
Share This Book
Module 9: Mendelian Genetics
Mendelian genetics, part 1: terminology.
Beginning students of biology always learn about Mendelian genetics. Inevitably, the study of inheritance always leads to additional questions. In fact, Mendelian inheritance patterns are exceedingly rare, especially in humans. We now know that inheritance is much more complex, usually involving many genes that interact in varied ways. Nonetheless, a clear understanding of basic inheritance patterns that follow Mendel’s original observations will provide a springboard for understanding current scientific exploration.
Inheritance patterns that follow Mendelian rules are as follows:
- Traits are governed by single genes
- There are two alternate forms of a gene, known as alleles
- Alleles are expressed as dominant and recessive
It just so happened that the traits Gregor Mendel observed in his pea plants did indeed conform to these rules. After collecting and analyzing his data, Gregor Mendel developed 2 laws of inheritance: The Law of Segregation and the Law of Independent Assortment.
- The Law of Segregation
- The Law of Independent Assortment
- Heterozygous
Part 2: Mendel’s First Law: Law of Segregation
The Law of Segregation states that alternative alleles of a trait segregate independently during meiosis.

Using a technique known as Punnett Square analysis, we will see how Mendel analyzed his monohybrid crosses to come up with the Law of Segregation.
Carefully follow each step to create a Punnett square analysis. You can use these same general procedures to analyze every Punnett Square you do!
Problem: In pea plants, height is coded for by the “T” gene. The dominant allele (T) codes for the tall phenotype while the recessive allele (t) codes for the short phenotype. Make a cross between a true breeding tall pea plant and a true breeding short pea plant.
- What are the phenotypes of the parent plants? The parents are considered the P generation.
- Determine the genotypes of each parent plant.
- Imagine each parent goes through meiosis to produce gametes. List the genotype(s) of the possible gametes that each parent would produce.
- Create a Punnett square that displays the genotypes of the possible offspring. Also label the phenotypes of the possible offspring. These offspring are considered the F 1 (first filial) generation.
- Now allow the F 1 generation to self-pollinate. What are the possible gametes that each F 1 parent can produce?
- Create a Punnett square that displays the genotypes of the possible offspring. Also label the phenotypes of the possible offspring. These offspring are considered the F 2 (second filial) generation.
Note: Always reduce the phenotypic and genotypic ratios to their lowest terms.
- What is the phenotypic ratio of the F 1 generation?
- What is the genotypic ratio of the F 1 generation?
- What is the phenotypic ratio of the F 2 generation?
- What is the genotypic ratio of the F 2 generation?
Part 3: Probability
Do the expected and observed phenotypic and genotypic ratios always match up in real life? In the case of flipping coins, we would expect to see heads 50% of the time and tails 50% of the time. But, does this always occur? Let’s explore!
- Working with a partner, take two coins and assume that heads represent the dominant allele (A) and tails represents the recessive allele (a). The genotype for each coin is heterozygous (Aa).
- Assume that each coin represents one parent. When a single coin is flipped, one gamete is formed (through the process of meiosis). If the flipped coin is on heads, then the gamete has the dominant allele (A). When both coins are flipped simultaneously, there will be two possible gametes that can combine through fertilization to form a zygote. Each time you flip both coins, you will record the “genotype” of the offspring.
| Expected results (after 100 flips) | Your results (# of flips with each outcome) | Class results | ||||
|---|---|---|---|---|---|---|
| Genotype | Expected count | Ratio (4 × count total flips) | Observed Count | Ratio | Observed count | Ratio |
| AA | ||||||
| Aa | ||||||
| aa | ||||||
| Total flips | 100 | 100 | ||||
- Report your results to your instructor so they can be included in the “Class Totals” column.
- What is the expected genotypic ratio for a cross between two Aa coins?
Think about It
Did the observed and expected genotypic ratios match? Why or why not?
Part 4: The Law of Independent Assortment
The Law of Independent Assortment states that genes located on different chromosomes assort independently from one another. To see the effects of this law, you must examine two different genes that are carried on two different chromosomes. We can investigate this phenomenon by looking at baby.
For this experiment, each group will examine a special ear of corn. These ears were created when a mama and papa corn plant, both heterozygous for seed color (P = purple, p= yellow) and seed shape (S = smooth, s = wrinkled), made baby corns. Corn is cool, because an ear of corn is just a whole bunch of babies held in one place! By counting the corn babies (each kernel is a baby), you can investigate the principle of Independent Assortment.
1 ear of corn/group
- What were the phenotypes of the mama and papa corn plants that gave rise to your cob of babes? (Read the previous paragraph to answer this question!)
- What were the genotypes of the mama and papa corn plants that gave rise to your cob of babes?
- What were all the possible gametes each parent corn could produce?
- Make a dihybrid cross illustrating all the possible baby corns produced by these parents. Then calculate the expected phenotypic ratios of the babies.
- Count and record the phenotypes of 100 kernels on your cob. Record your results below. ( Download the chart here. )
| Phenotypes of possible offspring | ||||
|---|---|---|---|---|
| Total Smooth Purple | Total Smooth Yellow | Total Wrinkled Purple | Total Wrinkled Yellow | |
| Expected ratio | ||||
| Total number counted | ||||
| Observed ratio [latex]\frac{(16\times\text{count})}{\text{total flips}}[/latex] | ||||
| Class observed ratio (class average) | ||||
Calculate the ratios using this formula:
[latex]\displaystyle\text{Phenotypic Ratio}=\frac{\text{number of possible combinations (16)}\times\text{number of kernels of a given phenotype}}{\text{total number of kernels counted (100)}}[/latex]
Note: If calculating class totals, the denominator in this equation is equal to the total of all kernels counted by all students in the class.
Lab Questions
- Do the observed phenotypes agree with the expected phenotypes? Why or why not?
- Can you determine the genotypes of the purple kernels or the smooth kernels in this lab exercise? Why or why not?
- Can you determine the genotypes of the yellow kernels or the wrinkled kernels in this lab exercise? Why or why not?
- Biology Labs . Authored by : Wendy Riggs . Provided by : College of the Redwoods . Located at : http://www.redwoods.edu . License : CC BY: Attribution
- Biology Article
Mendelian Genetics

Mendelian Genetics is a kind of biological inheritance that highlights the laws proposed by Gregor Mendel in 1866 and rediscovered in 1900. These laws faced a few controversies initially but when Mendel’s theories got integrated with the chromosome theory of inheritance, they soon became the heart of classical genetics. Later, Ronald Fisher combined these ideas with the theory of natural selection and forms a base for population genetics and modern evolutionary synthesis.
Table of Contents
Mendel’s experiments, mendel’s laws of inheritance, law of segregation, law of independent assortment, law of dominance.
Gregor Mendel performed breeding experiments in his garden to analyse patterns of inheritance. He opted for cross-bred normal pea plants with selective traits over various generations. When two plants were crossed that differed in a single trait (round peas vs. wrinkled seeds, short stems vs. tall stems, white flowers vs. purple flowers, etc), Mendel found that the next generation, F1 comprised of whole individuals that exhibit only one trait. However, after the generation was interbred, its offspring which is the F2 generation showed a 3:1 ratio wherein three individuals had similar traits like a parent.
Mendel theorized that genes could be formed by three possible combinations of heredity units that are said to be factors: AA, aa, Aa. The big ‘A’ shows the dominant factor and the small ‘a’ shows the recessive factor. The beginning plants were homozygous AA or aa, F1 generation was Aa and F2 generation was AA, aa or Aa. The interaction between these two finds the physical trait that is visible.
According to Mendel’s law of Dominance, when two organisms of separate traits are crossed, every offspring shows the trait of only one dominant character. The recessive trait is expressed phenotypically only if both factors are recessive.
Also Read: Non-Mendelian Inheritance
Mendel’s conclusions could be described in the following principles:
According to the law of segregation , every parent’s pair of genes or alleles divide and a single gene passes from every parent to an offspring. Which particular gene passes on in a pair is entirely up to chance.
According to the law of Independent Assortment, discrete pairs of alleles pass onto the offspring without depending on one another. Hence, the inheritance of genes at a particular region in a genome does not affect the inheritance of genes in a different region.
According to the law of dominance, recessive alleles are always masked by dominant alleles. Hence, a cross between a homozygous recessive and a homozygous dominant shows the dominant phenotype by still having a heterozygous genotype. This law could be explained by the monohybrid cross experiment. In the case of a cross among the two organisms with contrasting traits, the character that is visible in the F1 generation is known as dominant and the one that is suppressed is known as recessive. Every character is handled by a pair of dissimilar factors and only one among the characters shows the results. Please note that the law of dominance is true but not applicable from a global perspective.
- Endangered Species
Frequently Asked Questions – FAQs
What are the 3 principles of mendelian genetics, which allele is dominant and which one is recessive and on what basis is it decided, why did mendel choose garden peas for his experiments.

Put your understanding of this concept to test by answering a few MCQs. Click ‘Start Quiz’ to begin!
Select the correct answer and click on the “Finish” button Check your score and answers at the end of the quiz
Visit BYJU’S for all Biology related queries and study materials
Your result is as below
Request OTP on Voice Call
| BIOLOGY Related Links | |
Leave a Comment Cancel reply
Your Mobile number and Email id will not be published. Required fields are marked *
Post My Comment
Register with BYJU'S & Download Free PDFs
Register with byju's & watch live videos.

Snapsolve any problem by taking a picture. Try it in the Numerade app?
Concepts of Genetics
William s. klug, michael r. cummings, charlotte a. spencer, mendelian genetics - all with video answers.

Chapter Questions
In this chapter, we focused on the Mendelian postulates, probability, and pedigree analysis. We also considered some of the methods and reasoning by which these ideas, concepts, and techniques were developed. On the basis of these discussions, what answers would you propose to the following questions: (a) How was Mendel able to derive postulates concerning the behavior of "unit factors" during gamete formation, when he could not directly observe them? (b) How do we know whether an organism expressing a dominant trait is homozygous or heterozygous? (c) In analyzing genetic data, how do we know whether deviation from the expected ratio is due to chance rather than to another, independent factor? (d) since experimental crosses are not performed in humans, how do we know how traits are inherited?

Review the Chapter Concepts list on $\mathrm{p}$. 74 . The first five concepts provide a modern interpretation of Mendelian postulates. Based on these concepts, write a short essay that correlates Mendel's four postulates with what is now known about genes, alleles, and homologous chromosomes.

Albinism in humans is inherited as a simple recessive trait. For the following families, determine the genotypes of the parents and offspring. (When two alternative genotypes are possible, list both.) (a) Two normal parents have five children, four normal and one albino. (b) A normal male and an albino female have six children, all normal. (c) A normal male and an albino female have six children, three normal and three albino. (d) Construct a pedigree of the families in (b) and (c). Assume that one of the normal children in (b) and one of the albino children in (c) become the parents of eight children. Add these children to the pedigree, predicting their phenotypes (normal or albino).

Which of Mendel's postulates are illustrated by the pedigree that you constructed in Problem 3 ? List and define these postulates.
Discuss how Mendel's monohybrid results served as the basis for all but one of his postulates. Which postulate was not based on these results? Why?

What advantages were provided by Mendel's choice of the garden pea in his experiments?

Mendel crossed peas having round seeds and yellow cotyledons (seed leaves) with peas having wrinkled seeds and green cotyledons. All the $F_{1}$ plants had round seeds with yellow cotyledons. Diagram this cross through the $\mathrm{F}_{2}$ generation, using both the Punnett square and forked-line, or branch diagram, methods.

Based on the preceding cross, what is the probability that an organism in the $\mathrm{F}_{2}$ generation will have round seeds and green cotyledons and be true breeding?

Which of Mendel's postulates can only be demonstrated in crosses involving at least two pairs of traits? State the postulate.
Assume that you have a garden and some pea plants have solid leaves and others have striped leaves. You conduct a series of crosses $[(a) \text { through }(e)]$ and obtain the results given in the table. Define gene symbols and give the possible genotypes of the parents of each cross.

What is the basis for homology among chromosomes?

Two organisms, $A A B B C C D D E E$ and aabbccddee, are mated to produce an $\mathrm{F}_{1}$ that is self-fertilized. If the capital letters represent dominant, independently assorting alleles: (a) How many different genotypes will occur in the $\mathrm{F}_{2}$ ? (b) What proportion of the $\mathrm{F}_{2}$ genotypes will be recessive for all five loci? (c) Would you change your answers to (a) and/or (b) if the initial cross occurred between $A A b b C C$ddee$\times$aaBBccDDEE parents? (d) Would you change your answers to (a) and/or (b) if the initial cross occurred between $A A B B C C D D E E \times$ aabbccddEE parents?

Albinism, lack of pigmentation in humans, results from an autosomal recessive gene (a). Two parents with normal pigmentation have an albino child. (a) What is the probability that their next child will be albino? (b) What is the probability that their next child will be an albino girl? (c) What is the probability that their next three children will be albino?

Dentinogenesis imperfecta is a rare, autosomal, dominantly inherited disease of the teeth that occurs in about one in 8000 people (Witkop 1957 ). The teeth are somewhat brown in color, and the crowns wear down rapidly. Assume that a male with dentinogenesis imperfecta and no family history of the disease marries a woman with normal teeth. What is the probability that (a) their first child will have dentinogenesis imperfecta? (b) their first two children will have dentinogenesis imperfecta? (c) their first child will be a girl with dentinogenesis imperfecta?

In a study of black guinea pigs and white guinea pigs, 100 black animals were crossed with 100 white animals, and each cross was carried to an $\mathrm{F}_{2}$ generation. In 94 of the crosses, all the $\mathrm{F}_{1}$ offspring were black and an $\mathrm{F}_{2}$ ratio of 3 black: 1 white was obtained. In the other 6 cases, half of the $\mathrm{F}_{1}$ animals were black and the other half were white. Why? Predict the results of crossing the black and white $\mathrm{F}_{1}$ guinea pigs from the 6 exceptional cases.
Mendel crossed peas having round green seeds with peas having wrinkled yellow seeds. All $\mathrm{F}_{1}$ plants had seeds that were round and yellow. Predict the results of testcrossing these $\mathrm{F}_{1}$ plants.
Thalassemia is an inherited anemic disorder in humans. Affected individuals exhibit either a minor anemia or a major anemia. Assuming that only a single gene pair and two alleles are involved in the inheritance of these conditions, is thalassemia a dominant or recessive disorder?

A certain type of congenital deafness in humans is caused by a rare autosomal (not X-linked) dominant gene. (a) In a mating involving a deaf man and a deaf woman (both heterozygous), would you expect all the children to be deaf? Explain your answer. (b) In a mating involving a deaf man and a deaf woman (both heterozygous), could all the children have normal hearing? Explain your answer. (c) Another form of deafness is caused by a rare autosomal recessive gene. In a mating involving a deaf man and a deaf woman, could some of the children have normal hearing? Explain your answer.

In assessing data that fell into two phenotypic classes, a geneticist observed values of $250: 150 .$ She decided to perform a $\chi^{2}$ analysis by using the following two different null hypotheses: (a) the data fit a 3: 1 ratio, and (b) the data fit a 1: 1 ratio. Calculate the $\chi^{2}$ values for each hypothesis. What can be concluded about each hypothesis?
The basis for rejecting any null hypothesis is arbitrary. The researcher can set more or less stringent standards by deciding to raise or lower the $p$ value used to reject or not reject the hypothesis. In the case of the chi-square analysis of genetic crosses, would the use of a standard of $p=0.10$ be more or less stringent about not rejecting the null hypothesis? Explain.

Among dogs, short hair is dominant to long hair and dark coat color is dominant to white (albino) coat color. Assume that these two coat traits are caused by independently segregating gene pairs. For each of the crosses given below, write the most probable genotype (or genotypes if more than one answer is possible for the parents. It is important that you select a realistic symbol set and define each symbol below. Assume that for cross (d), you were interested in determining whether fur color follows a 3: 1 ratio. Set up (but do not complete the calculations) a Chi-square test for these data [fur color in cross $(\mathrm{d})]$.

Draw all possible conclusions concerning the mode of inheritance of the trait portrayed in each of the following limited pedigrees. (Each of the four cases is based on a different trait.) a. b. c. d.
In a family of eight children, what is the probability that (a) the third child is a girl? (b) six of the children are boys? (c) all the children are girls? (d) there are four boys and four girls? Assume that the probability of having a boy is equal to the probability of having a girl $(p=1 / 2)$.
In a family of six children, where one grandparent on either side has red hair, what mathematical expression predicts the probability that two of the children have red hair?

The autosomal (not X-linked) gene for brachydactyly, short fingers, is dominant to normal finger length. Assume that a female with brachydactyly in the heterozygous condition is married to a man with normal fingers. What is the probability that (a) their first child will have brachydactyly? (b) their first two children will have brachydactyly? (c) their first child will be a brachydactylous girl?
Galactosemia is a rare recessive disorder caused by the deficiency of galactose- 1 -phosphate uridylyltransferase, leading to the accumulation of toxic levels of galactitol in the blood. It leads to a $75 \%$ mortality rate in infants as infants cannot metabolize galactose from breast milk. In many countries, newborns are given a heel prick test to measure the levels of metabolic enzymes. As a genetic counselor, how would you explain to a couple whose baby has tested positive for galactosemia where the disease has come from?

Two true-breeding pea plants were crossed. One parent is round, terminal, violet, constricted, while the other expresses the respective contrasting phenotypes of wrinkled, axial, white, full. The four pairs of contrasting traits are controlled by four genes, each located on a separate chromosome. In the $\mathrm{F}_{1}$ only round, axial, violet, and full were expressed. In the $\mathrm{F}_{2},$ all possible combinations of these traits were expressed in ratios consistent with Mendelian inheritance. (a) What conclusion about the inheritance of the traits can be drawn based on the $\mathrm{F}_{1}$ results? (b) In the $\mathrm{F}_{2}$ results, which phenotype appeared most frequently? Write a mathematical expression that predicts the probability of occurrence of this phenotype. (c) Which $\mathrm{F}_{2}$ phenotype is expected to occur least frequently? Write a mathematical expression that predicts this probability. (d) In the $F_{2}$ generation, how often is either of the $P_{1}$ phenotypes likely to occur? (e) If the $F_{1}$ plants were testcrossed, how many different phenotypes would be produced? How does this number compare with the number of different phenotypes in the $\mathrm{F}_{2}$ generation just discussed?
Tay-Sachs disease (TSD) is an inborn error of metabolism that results in death, often by the age of $2 .$ You are a genetic counselor interviewing a phenotypically normal couple who tell you the male had a female first cousin (on his father's side) who died from TSD and the female had a maternal uncle with TSD. There are no other known cases in either of the families, and none of the matings have been between related individuals. Assume that this trait is very rare. (a) Draw a pedigree of the families of this couple, showing the relevant individuals. (b) Calculate the probability that both the male and female are carriers for TSD. (c) What is the probability that neither of them is a carrier? (d) What is the probability that one of them is a carrier and the other is not? [Hint: The $p$ values in (b), (c), and (d) should equal $1 .]$
Datura stramonium (the Jimsonweed) expresses flower colors of purple and white and pod textures of smooth and spiny. The results of two crosses in which the parents were not necessarily true breeding are shown at the top of the next column. (a) Based on these results, put forward a hypothesis for the inheritance of the purple/white and smooth/spiny traits. (b) Assuming that true-breeding strains of all combinations of traits are available, what single cross could you execute and carry to an $\mathrm{F}_{2}$ generation that will prove or disprove your hypothesis? Assuming your hypothesis is correct, what results of this cross will support it??
The wild-type (normal) fruit fly, Drosophila melanogaster, has straight wings and long bristles. Mutant strains have been isolated that have either curled wings or short bristles. The genes representing these two mutant traits are located on separate chromosomes. Carefully examine the data from the five crosses shown on the top of the following page (running across both columns). (a) Identify each mutation as either dominant or recessive. In each case, indicate which crosses support your answer. (b) Assign gene symbols and, for each cross, determine the genotypes of the parents.

An alternative to using the expanded binomial equation and Pascal's triangle in determining probabilities of phenotypes in a subsequent generation when the parents' genotypes are known is to use the following equation: $\frac{n !}{s ! t !} a^{s} b^{t}$ where $n$ is the total number of offspring, $s$ is the number of offspring in one phenotypic category, $t$ is the number of offspring in the other phenotypic category, $a$ is the probability of occurrence of the first phenotype, and $b$ is the probability of the second phenotype. Using this equation, determine the probability of a family of 5 offspring having exactly 2 children afflicted with sickle-cell anemia (an autosomal recessive disease $)$ when both parents are heterozygous for the sickle-cell allele.

To assess Mendel's law of segregation using tomatoes, a truebreeding tall variety (SS) is crossed with a true-breeding short variety $(s s) .$ The heterozygous $F_{1}$ tall plants $(S s)$ were crossed to produce two sets of $\mathrm{F}_{2}$ data, as follows. $\begin{array}{cc}\text { Set I } & \text { Set II } \\ 30 \text { tall } & 300 \text { tall } \\ 5 \text { short } & 50 \text { short }\end{array}$ (a) Using the $\chi^{2}$ test, analyze the results for both datasets. Calculate $\chi^{2}$ values and estimate the $p$ values in both cases. (b) From the above analysis, what can you conclude about the importance of generating large datasets in experimental conditions?
Albinism, caused by a mutational disruption in melanin (skin pigment production, has been observed in many species, including humans. In 1991 , the only documented observation of an albino humpback whale (named "Migaloo") was observed near New South Wales. Recently, Polanowski and coworkers (Polanowski, A., S. Robinson-Laverick, and D. Paton. 2012. Journal of Heredity $103: 130-133$ ) studied the genetics of humpback whales from the east coast of Australia, including Migaloo. (a) Do you think that Migaloo's albinism is more likely caused by a dominant or recessive mutation? Explain your reasoning. (b) What data would be helpful in determining the answer to part (a)?

(a) Assuming that Migaloo's albinism is caused by a rare recessive gene, what would be the likelihood of the establishment of a natural robust subpopulation of albino white humpback whales in this population? (b) Assuming that Migaloo's albinism is caused by a rare dominant gene, what would be the likelihood of the establishment of a natural robust subpopulation of albino white humpback whales in this population?

Assume that Migaloo's albinism is caused by a rare recessive gene. (a) In a mating of two heterozygous, normally pigmented whales, what is the probability that the first three offspring will all have normal pigmentation? (b) What is the probability that the first female offspring is normally pigmented? (c) What is the probability that the first offspring is a normally pigmented female?
Dentinogenesis imperfecta is a tooth disorder involving the production of dentin sialophosphoprotein, a bone-like component of the protective middle layer of teeth. The trait is inherited as an autosomal dominant allele located on chromosome 4 in humans and occurs in about 1 in 6000 to 8000 people. Assume that a man with dentinogenesis imperfecta, whose father had the disease but whose mother had normal teeth, married a woman with normal teeth. They have six children. What is the probability that their first child will be a male with dentinogenesis imperfecta? What is the probability that three of their six chil- dren will have the disease?
- History & Society
- Science & Tech
- Biographies
- Animals & Nature
- Geography & Travel
- Arts & Culture
- Games & Quizzes
- On This Day
- One Good Fact
- New Articles
- Lifestyles & Social Issues
- Philosophy & Religion
- Politics, Law & Government
- World History
- Health & Medicine
- Browse Biographies
- Birds, Reptiles & Other Vertebrates
- Bugs, Mollusks & Other Invertebrates
- Environment
- Fossils & Geologic Time
- Entertainment & Pop Culture
- Sports & Recreation
- Visual Arts
- Demystified
- Image Galleries
- Infographics
- Top Questions
- Britannica Kids
- Saving Earth
- Space Next 50
- Student Center

- Why is Gregor Mendel famous?

Mendelian inheritance
Our editors will review what you’ve submitted and determine whether to revise the article.
- Roger Williams University Pressbooks - Introduction to Molecular and Cell Biology - Mendelian Genetics
- Biology LibreTexts - Mendelian inheritance
- National Center for Biotechnology Information - PubMed Central - Mendelian Genetics
- Thompson Rivers University - Human Biology - Mendelian Inheritance
- North Dakota State University - Mendel's First Law of Genetics

Mendelian inheritance , the principles of heredity formulated by Austrian-born botanist, teacher, and Augustinian prelate Gregor Mendel in 1865. These principles compose what is known as the system of particulate inheritance by units, or genes . The later discovery of chromosomes as the carriers of genetic units supported Mendel’s two basic laws, known as the law of segregation and the law of independent assortment .

In modern terms, the first of Mendel’s laws states that genes are transferred as separate and distinct units from one generation to the next. The two members ( alleles ) of a gene pair, one on each of paired chromosomes, separate during the formation of sex cells by a parent organism. One-half of the sex cells will have one form of the gene, one-half the other form; the offspring that result from these sex cells will reflect those proportions.

A modern formulation of the second law, the law of independent assortment, is that the alleles of a gene pair located on one pair of chromosomes are inherited independently of the alleles of a gene pair located on another chromosome pair and that the sex cells containing various assortments of these genes fuse at random with the sex cells produced by the other parent.
Mendel also developed the law of dominance , in which one allele exerts greater influence than the other on the same inherited character . Mendel developed the concept of dominance from his experiments with plants, based on the supposition that each plant carried two trait units, one of which dominated the other. For example, if a pea plant with the alleles T and t ( T = tallness, t = shortness) is equal in height to a T T individual, the T allele (and the trait of tallness) is completely dominant. If the T t individual is shorter than the T T but still taller than the t t individual, T is partially or incompletely dominant—i.e., it has a greater influence than t but does not completely mask the presence of t , which is recessive.
- BiologyDiscussion.com
- Follow Us On:
- Google Plus
- Publish Now
Essay on the Mendel’s Law of Inheritance | Genetics

ADVERTISEMENTS:
In this essay we will discuss about:- 1. Law’s of Hereditary 2. Reasons for Mendel’s Success 3. Reasons for Overlooking of Mendel’s Results 4. Extensions of Mendelian Concepts.
Essay on the Law’s of Hereditary:
Mendel laid the foundation of the science of genetics through the discovery of basic principles of heredity. He conducted his experiments with garden pea (pisum sativum) in a small monastery garden for over seven years (1856-1864) and discovered two important laws of heredity, viz., 1. law of segregation, and 2. law of independent assortment. These are briefly presented below.
I. Law of Segregation :
This law states that alleles segregate or separate from each other during gamete formation and pass on to different gametes in equal number. In other words, when alleles for two contrasting characters come together in a hybrid, they do not blend, contaminate or affect each other while together.
The different genes separate from each other in a pure form, pass on to different gametes formed by a hybrid and then go to different individuals in the offspring of the hybrid.
Thus main features of this law are as follows:
i. When a dominant and a recessive allele of a gene come together in a hybrid after crossing between two plants having contrasting characters, they do not mix or blend together.
ii. They remain together in pure form without affecting each other. For this reason, law of segregation is also known as law of purity of gametes.
iii. They separate into different gametes in equal number. Each gamete has only one type of allele (say either A or a).
iv. Separation of two alleles of a gene during gamete formation takes place usually due to the separation of homologous chromosomes during meiosis (anaphase 1), because alleles are located in the chromosomes.
v. With complete dominance, segregation leads to phenotypic ratio of 3 : 1 in F 2 for characters governed by single gene, and 9:3:3:1 ratio for characters controlled by two genes.
vi. If crossing over does not take place, segregation of genes takes place during anaphase I. If crossing over occurs, segregation of genes will take place during anaphase II.
When we make a cross between red (RR) and white (rr) flowered plants, we get red colour of flower in F 1 . In the F 1 both the alleles R and r remain together without blending or mixing with each other, though only the effect of dominant allele is visible. In F 2 , allele for red flower colour and white flower colour segregate during gamete formation and pass on to the gametes in equal number.
Thus two types of gametes, viz., R and r are formed. Each gamete has either R or r allele. When the F 1 is self-pollinated, individuals with three genotypes, viz., RR, Rr and rr are obtained in F 2 . Here RR and Rr are all red and only rr individuals are white (Fig. 7.1). Thus a phenotypic ratio of 3 red: 1 white is obtained. The overall mechanism is represented below.

When selfed seeds of RR were grown in F 3 , they all produced all the true breeding individuals for red flower colour. The Rr individuals showed segregation in F 3 similar to segregation in F 2 generation. Individuals with rr genotypes were found true breeding for white flower colour when their selfed seeds were raised in F 3 generation.
II. Law of Independent Assortment :
This is the second law of inheritance discovered by Mendel. This law states that when two pairs of gene enter in F 1 combination, both of them have their independent dominant effect. These genes segregate when gametes are formed, but the assortment occurs randomly and quite freely.
Thus main features of this law are given below:
i. This law explains simultaneous inheritance of two plant characters.
ii. In F 1 when two genes controlling two different characters, come together, each gene exhibits independent dominant behaviour without affecting or modifying the effect of other gene.
iii. These gene pairs segregate during gamete formation independently.
iv. The alleles of one gene can freely combine with the alleles of another gene. Thus each allele of one gene has an equal chance to combine with each allele of another gene.
v. Each of the two gene pairs when considered separately, exhibits typical 3 : 1 segregation ratio in F 2 generation. This is a typical monohybrid segregation ratio.
vi. Random or free assortment of alleles of two genes leads to formation of new gene combinations.
When plants of garden pea with yellow round seeds are crossed with plants having green wrinkled seeds, we get yellow round seeds in F 1 . Thus yellow colour of seed exhibits dominance over green and round seeds shape over wrinkled independently.
The F 1 produces four types of gametes, viz., yellow round (YR), yellow wrinkled (Yr), green round (yR), and green wrinkled (yr). Selfing of F 1 gives rise to all above four types of individuals in 9 : 3 : 3 : 1 ratio (Fig. 7.2).
The behaviour of all these genotypes was studied in F 3 generation. Out of nine yellow round individuals only one (YYRR) was found true breeding in F 3 generation. The other eight individuals showed segregation of various types.
Similarly, out of 3 yellow wrinkled individuals only one (YYrr) bred true and others segregated in 3 : 1 ratio. Same thing happened with green round individuals. The green wrinkled individual was also true breeding (Table 7.2).

Reasons for Mendel’s Success :
Investigations to unravel the mechanism of inheritance were made by several workers. However, only Mendel could get success in explaining the laws of inheritance.
The important factors which are responsible for Mendel’s success are briefly described below:
I. Proper Maintenance of Records :
Mendel had a systematic record of all the observations which he recorded on various characters in different generations. This helped him in proper understanding of laws responsible for the transmission of characters from one generation to other.
II. Study of Individual Character :
Mendel laid emphasis on the study of individual character, which helped him in systematic analysis of various characters. He studied seven characters of garden pea. Other workers studied the individual as a whole which confused the whole issue and they could not come up with concrete results.
III. Choice of Material :
Mendel selected garden pea for his investigations. Garden pea is a hermaphrodite, self-fertilized and short duration crop. This helped Mendel in maintaining the purity of material and raising more than one generation in a year. Moreover, flowers of garden pea are also ideal for hand emasculation and pollination.
IV. Maintenance of Purity :
Mendel always used pure breeding parents or lines for hybridization, which helped him in studying the inheritance of individual character in a systematic way. He used to maintain purity by growing different lines in separate plots to avoid mechanical admixture or contamination. Use of heterozygous material poses several difficulties in drawing general conclusions.
V. Knowledge of Shortfalls of Earlier Workers :
Mendel systematically recorded the reasons for the failure of earlier workers in investigating the mechanism of inheritance. This helped Mendel to think in new direction and better planning of his work.
VI. Mathematical Background :
Mendel had very good background of physics and mathematics. He studied mathematics at the University of Vienna where he was sent for higher studies by the monastery. His mathematical knowledge proved boon for him, which helped Mendel in explaining the segregation of characters in F 2 and F 3 generations in terms of segregation ratios.
He was able to understand that in natural population, it is very difficult to get segregation perfectly in a particular ratio. A slight deviation from the exact ratio will generally be observed.
VII. Proper Choice of Characters :
In garden pea, Mendel could easily select contrasting (opposite) forms for various characters, which helped him in getting clear-cut groups in F 2 and F 3 generations. Moreover, all the seven characters which Mendel studied in garden pea, were qualitative in nature.
Such characters always display discontinuous variation, which permits classification of individuals into different clear-cut groups for these characters. This also helped Mendel in generalizing his results.
Reasons for Overlooking of Mendel’s Results :
Mendel presented results of his seven years hard and devoted work in the meetings of the Natural History Society of Brunn on 8th February and 8th March, 1865 in two papers. His papers on “Experiments in Plants” were published in detail in the annual proceedings of the society in 1866 in German language.
These proceedings were distributed to several libraries in Europe and America. However, Mendel’s work remained overlooked for 34 years. Mendel died in 1884 and his work came to light 16 years after his death when three different scientists viz., de Vries (Holland), Correns (Germany) and Tschermak (Austria) independently reached at the same conclusions in 1900 which were drawn by Mendel 34 years ago, i.e. in 1866.
The important reasons for the neglect of Mendel’s revolutionary findings related to mechanism of inheritance for such a long time are briefly presented below:
i. Mendel generalized his results based on his studies on garden pea. Later on he worked on hawkweed (Hieraceum) on the advice of C.V. Nageli. Mendel could not prove his results on this plant as the embryo is formed from the ovule without fertilization (diploid parthenogenesis). This created doubt in the mind of scientists about the results of Mendel.
ii. Since Mendel has a good background of mathematics, he explained his results with the help on mathematics. The scientists at that time did not appreciate this approach.
iii. Mendel could not support his findings through cytological investigations as cytological studies were not well developed at that time.
iv. After his failure to demonstrate the results on hawkweed, Mendel lost interest in research work and devoted most of his time with the work of monastery. Moreover, he did not give proper publicity to his work and kept quiet.
Extensions of Mendelian Concepts :
The basic principles of heredity were initially discovered by Mendel in 1866 and rediscovered by de Vries, Correns and Tschermak in 1900. Later on these principles were clarified and confirmed by several researchers and some new concepts were investigated. Some of the new concepts were at variance with the findings of Mendel.
These are called as Mendelian deviations or exceptions or anomalies.
Such investigations include:
i. Incomplete dominance,
ii. Co-dominance,
iii. Multiple alleles,
iv. Linkage,
v. Lethal genes,
vi. Gene interactions,
vii. Pleiotropic gene effect,
viii. Polygenes,
ix. Environmental effects, and
x. Cytoplasmic or maternal effects.
Mendel did not come across these findings. In fact these are extensions of Mendelian concepts. A brief description of these concepts in presented below.
I. Incomplete Dominance :
Mendel always observed complete dominance of one allele over the other for all the seven characters which he studied in garden pea. Later on cases of incomplete dominance were reported. For example, in four ‘o’ clock plant (Mirabilis jalapa) there are two types of flowers, viz., red and white.
A cross between red and white flowered plants produced plants with intermediate flower colour, i.e., pink colour in F 1 and a modified ratio of 1 red : 2 pink : 1 white was observed in F 2 (Fig. 7.3).

II. Co-Dominance :
In case of co-dominance both alleles express their phenotypes in heterozygote. The example is AB blood group in human. The people who have blood type AB are heterozygous exhibiting phenotypes for both the I A and I B alleles. In other words, heterozygotes for co-dominant alleles are phenotypically similar to both parental types.
The main difference between co-dominance and incomplete dominance lies in the way in which genes act. In case of co-dominance, both alleles are active, while in case of incomplete dominance only one allele (dominant) is active.
III. Multiple Alleles :
Mendel always observed two allelic forms of a gene. Now cases are known where a gene has more than two allelic forms, although only two can exist in a diploid cell at a time. Existence of more than two alleles for a gene is called multiple alleles. Examples of multiple alleles are ABO blood group alleles in human, coat colour in rabbit and self-incompatibility alleles in tobacco.
IV. Linkage :
Mendel always observed independent assortment of genes. Later on cases of linkage were reported by Bateson and Punnett in 1906 in pea, Hutchinson in maize and Morgan (1910) in Drosophila. In a di-hybrid test cross, they observed higher frequencies of parental types than recombinants instead of 1 : 1 : 1 : 1 ratio. This led to modification of the concept of independent assortment.
V. Lethal Genes :
Gene which causes the death of its earner when in homozygous condition is called lethal gene. Mendel’s findings were based on equal survival of all genotypes. In the presence of lethal genes, the normal segregation ratio of 3 : 1 is modified into 2 : 1 ratio. Lethal genes have been reported in both animals as well as plants. In mice, allele for yellow coat colour is dominant over grey.
When a cross is made between yellow and grey, a ratio of 1 : 1 for yellow and grey mice was observed. This indicated that yellow mice are always heterozygous, because yellow homozygotes are never born because of homozygous lethality. Such genes were not observed by Mendel. He always got 3 : 1 ratio in F 2 for single gene characters.
VI. Gene Interactions :
When the expression of an allele of one gene pair depends on the presence of a specific allele of another pair, it is known as gene interaction. Mendel observed 9:3:3:1 ratio in F 2 from a dihybrid cross. Later on many deviations of this phenotypic ratio were observed in dihybrid crosses. The modified ratios included 9:7, 9:3:4, 12:3: 1, 13:3: 15: 1 and 9:6:1 in different crop plants.
VII. Pleiotropic Gene Effects :
Mendel observed that one gene controls the expression of only one character. Later on cases were observed in which one gene was found to govern the expression of two or more characters. Example is white eye allele in Drosophila. This allele affects eye colour, shape of spermathica, fecundity and testicular membrane.
VIII. Polygenes :
Mendel always observed that each character is governed by a single gene. Later on Nilsson Ehle observed that some characters are controlled by several genes and each of such gene has additive effect in the expression of character. This concept led to the foundation of polygenic inheritance.
East (1916) demonstrated that polygenic characters were perfectly in agreement with Mendelian segregation and later on Fisher and Wright provided a mathematical basis for the genetic interpretation of polygenic characters.
IX. Environmental Effects :
Genes can interact not only with other genes but also with the environment to produce the final phenotype. Thus phenotype is the result of the interaction between genotype and environment. It leads directly to the concept of penetrance and expressivity. The importance of environment was first realised by Johannsen. He coined the terms genotype and phenotype.
X. Maternal Effects :
Mendel did not observe any difference between direct and reciprocal crosses. Later investigations revealed the presence of significant difference in the reciprocal crosses, which led to the concept of cytoplasmic inheritance.
Related Articles:
- Mendel’s Law of Inheritance | Genetics
- Mendel’s Postulates and Laws of Inheritance (With Diagram) | Botany
Biology , Genetics , Mendel’s Law of Inheritance
- Anybody can ask a question
- Anybody can answer
- The best answers are voted up and rise to the top
Forum Categories
- Animal Kingdom
- Biodiversity
- Biological Classification
- Biology An Introduction 11
- Biology An Introduction
- Biology in Human Welfare 175
- Biomolecules
- Biotechnology 43
- Body Fluids and Circulation
- Breathing and Exchange of Gases
- Cell- Structure and Function
- Chemical Coordination
- Digestion and Absorption
- Diversity in the Living World 125
- Environmental Issues
- Excretory System
- Flowering Plants
- Food Production
- Genetics and Evolution 110
- Human Health and Diseases
- Human Physiology 242
- Human Reproduction
- Immune System
- Living World
- Locomotion and Movement
- Microbes in Human Welfare
- Mineral Nutrition
- Molecualr Basis of Inheritance
- Neural Coordination
- Organisms and Population
- Photosynthesis
- Plant Growth and Development
- Plant Kingdom
- Plant Physiology 261
- Principles and Processes
- Principles of Inheritance and Variation
- Reproduction 245
- Reproduction in Animals
- Reproduction in Flowering Plants
- Reproduction in Organisms
- Reproductive Health
- Respiration
- Structural Organisation in Animals
- Transport in Plants
- Trending 14
Privacy Overview
| Cookie | Duration | Description |
|---|---|---|
| cookielawinfo-checkbox-analytics | 11 months | This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Analytics". |
| cookielawinfo-checkbox-functional | 11 months | The cookie is set by GDPR cookie consent to record the user consent for the cookies in the category "Functional". |
| cookielawinfo-checkbox-necessary | 11 months | This cookie is set by GDPR Cookie Consent plugin. The cookies is used to store the user consent for the cookies in the category "Necessary". |
| cookielawinfo-checkbox-others | 11 months | This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Other. |
| cookielawinfo-checkbox-performance | 11 months | This cookie is set by GDPR Cookie Consent plugin. The cookie is used to store the user consent for the cookies in the category "Performance". |
| viewed_cookie_policy | 11 months | The cookie is set by the GDPR Cookie Consent plugin and is used to store whether or not user has consented to the use of cookies. It does not store any personal data. |
IMAGES
VIDEO
COMMENTS
7. A woman has a daughter. There are three men whom she claims might have been the father of the child. The judge in the paternity court orders that all three men, the child, and the mother have blood tests. The results are: mother, Type A; Daughter, Type O; Man #1, Type AB; Man #2, Type B; Man #3, Type O.
Test cross. Study Guide Questions. Understand Gregor Mendel's experiments, his results, and his conclusions. Clearly relate MEIOSIS to Mendel's work. Given data from a genetic cross, be able to determine information about how the trait in question is inherited. Be able to successfully "do" both monohybrid and dihybrid crosses.
Khanmigo is now free for all US educators! Plan lessons, develop exit tickets, and so much more with our AI teaching assistant.
Mendel's experiments have given true data that proves the laws to be true. Our experiments included that same steps, so they had the same significance. There are two specific hypotheses that come along with this experiment. 1. The experiment using the mung bean plants will follow the principle of segregation. 2.
9. Two corn homozygous, white-seeded corn lines were crossed and all progeny were red-seeded. These red-seeded F 1 progeny were selfed, and the population segregated 9 red-seeded:7 white-seeded. Explain these results by determining the number of genes controlling seed coat colors and giving the genotypes of the parents. 10.
By experimenting with pea plant breeding, Mendel developed three principles of inheritance that described the transmission of genetic traits, before anyone knew genes existed. Mendel's insight ...
Mendel's laws (principles) of segregation and independent assortment are both explained by the physical behavior of chromosomes during meiosis. Segregation occurs because each gamete inherits only one copy of each chromosome. Each chromosome has only one copy of each gene; therefore each gamete only gets one allele.
Introduction. Figure 18.2 Johann Gregor Mendel is considered to be the father of genetics. Genetics is the study of heredity. Johann Gregor Mendel (1822-1884) set the framework for genetics long before chromosomes or genes had been identified, at a time when meiosis was not well understood (Figure 18.2). Mendel selected a simple biological ...
Part 1: Terminology. Beginning students of biology always learn about Mendelian genetics. Inevitably, the study of inheritance always leads to additional questions. In fact, Mendelian inheritance patterns are exceedingly rare, especially in humans. We now know that inheritance is much more complex, usually involving many genes that interact in ...
Mendelian Genetics. Mendelian Genetics is a kind of biological inheritance that highlights the laws proposed by Gregor Mendel in 1866 and rediscovered in 1900. These laws faced a few controversies initially but when Mendel's theories got integrated with the chromosome theory of inheritance, they soon became the heart of classical genetics.
Problem 1. In this chapter, we focused on the Mendelian postulates, probability, and pedigree analysis. We also considered some of the methods and reasoning by which these ideas, concepts, and techniques were developed. On the basis of these discussions, what answers would you propose to the following questions:
Study with Quizlet and memorize flashcards containing terms like Summarize Mendel's experiments, and the three laws of inheritance that make up the foundation of Mendelian Genetics., Create an example of a monohybrid cross. Write out a sample problem and the parents' genotypes. Solve a Punnett square and determine the genotypic and phenotypic ratios for your example., Create an example of a ...
BSC 2011MENDELIAN GENETICS PROBLEMSThe following problems are provided to develop your skill and test your understanding of solving pro. ems in the patterns of inheritance. They will be most h. pful if you solve them on your own. However, you should seek help if. u find you cannot answer a problem. Most of these problems are fairly simple, yet ...
Mendelian inheritance, the principles of heredity formulated by Austrian-born botanist, teacher, and Augustinian prelate Gregor Mendel in 1865. These principles compose what is known as the system of particulate inheritance by units, or genes. The later discovery of chromosomes as the carriers of genetic units supported Mendel's two basic ...
Study with Quizlet and memorize flashcards containing terms like When Mendel did crosses of true-breeding purple- and white-flowered pea plants, the white-flowered trait disappeared from the F1 generation but reappeared in the F2 generation. Use genetic terms to explain why that happened, What pattern(s) of inheritance is/are demonstrated by the ABO blood group alleles?, You are a licensed ...
Terms in this set (33) Mendelian genetics. provides basic understanding for how regulated occurrence of variation (+natural selection) can lead to new species. parental generation. P; original plants involved in making the crosses. first filial. F₁; offspring of first cross of P generation. second filial. F₂; offspring of F₁ generation.
ADVERTISEMENTS: In this essay we will discuss about:- 1. Law's of Hereditary 2. Reasons for Mendel's Success 3. Reasons for Overlooking of Mendel's Results 4. Extensions of Mendelian Concepts. Essay on the Law's of Hereditary: Mendel laid the foundation of the science of genetics through the discovery of basic principles of heredity. He conducted his […]